Cuáles son las ventajas y desventajas del fenómeno del hermafroditismo. Hermafroditismo y sus tipos. Quejas y anamnesis.
¿Cuáles son las ventajas del hermafroditismo y las características de la dioica?
El hermafroditismo existe porque en algunos casos se observa. Baja densidad de población
La dioecia tiene la ventaja de que no se produce homocigotización de mutaciones, por lo que hay menos muertes durante la reproducción. La variabilidad genética de la población está aumentando.
Aromorfosis de plantas y animales que contribuyen a la conquista de la tierra:
Plantas: Tejidos tegumentarios (epidermis, corcho), tejidos conductores, tejidos mecánicos, proceso sexual independiente del goteo de agua líquida,
Animales: aparición de pulmones, cubiertas corporales primero húmedas, luego secas y duras, desarrollo de órganos sensoriales adaptados a las condiciones terrestres (visión), reproducción independiente del agua, mejora del sistema excretor (los productos de excreción se volvieron completamente diferentes debido a la falta de agua) .
Seminario No. 6
Conceptos elementales de la teoría sintética de la emisión.
1. Propiedades básicas de la materia viva de nuestro planeta.
Es necesario tener una comprensión general del objeto de la evolución, las propiedades básicas de los seres vivos. Por otra parte, propiedades como el metabolismo, la movilidad, la irritabilidad, el crecimiento, la reproducción y la adaptabilidad también se encuentran en la naturaleza inanimada y, por lo tanto, no pueden considerarse propiedades específicas de los vivos.
Cinco axiomas de la biología teórica. En uno de los últimos y más exitosos intentos, los seres vivos se caracterizan por los siguientes rasgos, formulados por B. M. Mednikov (1982) en forma de axiomas de biología teórica:
1) Todos los organismos vivos resultan ser una unidad de un fenotipo y un programa para su construcción (genotipo), transmitido por herencia de generación en generación (axioma de A. Weisman).
2) El programa genético se forma de forma matricial. El gen de la generación anterior se utiliza como matriz sobre la cual se construye el gen de la generación futura (axioma de N.K. Koltsov).
3) En el proceso de transmisión de generación en generación, los programas genéticos, como resultado de diversas razones, cambian de forma aleatoria y no dirigida, y sólo por casualidad tales cambios pueden tener éxito en un entorno determinado (primer axioma de Ch.D.a).
4) Los cambios aleatorios en los programas genéticos durante la formación de un fenotipo se amplifican muchas veces (el axioma de N.V. Timofeev-Resovsky).
5) Los cambios repetidamente mejorados en los programas genéticos están sujetos a la selección por las condiciones ambientales (segundo axioma de Charles Darwin).
Nekot. propiedades que están directamente relacionadas con el proceso de desarrollo evolutivo que ocurre en todas partes.
La discreción y la integridad son dos propiedades fundamentales de la organización de la vida en la Tierra.
Los objetos vivos en la naturaleza están relativamente aislados unos de otros (individuos, poblaciones, especies). Cualquier animal multicelular individual está formado por células, y cualquier célula y criatura unicelular está formado por ciertos orgánulos. Los orgánulos están formados por sustancias orgánicas discretas, generalmente de alto peso molecular. a su vez, están formados por átomos discretos, partículas elementales (¡también discretas!). Al mismo tiempo, una organización compleja es impensable sin la interacción de sus partes y estructuras, sin integridad. La integridad de los sistemas biológicos es cualitativamente diferente de la integridad de los seres no vivos, y principalmente en que la integridad de los vivos se mantiene en el proceso de desarrollo. Se caracterizan por la entropía negativa. Es probable que los seres vivos presenten la capacidad de autoorganizar la materia.
Reduplicación convariante ( la autorreproducción con cambios), realizada sobre la base del principio matricial (la suma de los tres primeros axiomas), es aparentemente la única propiedad específica de la vida (en la forma de existencia que conocemos en la Tierra). Se basa en la capacidad única de autorreproducción de los principales sistemas de control (ADN, cromosomas y genes). Cuando los sistemas de control se autorreproducen en los organismos vivos, no se produce una repetición mecánica, sino una reproducción con la introducción de cambios.
2. Niveles de organización de la vida. en el piso. Qué eventos evolutivos ocurren en cada nivel de la organización.
Si intentamos identificar los niveles principales que reflejan no tanto los niveles de estudio, sino más bien los niveles de organización de la vida en la Tierra, entonces los principales criterios para dicha identificación deberían ser Se reconoce la presencia de estructuras elementales, discretas y fenómenos elementales específicos. (Se destacó a N.V. Timofeev-Resovsky y otros).
Nivel genético molecular. Las unidades elementales en este nivel son los sistemas de control básicos (ADN, cromosomas y genes). Los principales fenómenos elementales asociados con ellos pueden considerarse su capacidad para sufrir reduplicación convariante, cambios estructurales locales (mutaciones) y la capacidad de transmitir información almacenada en ellos a los sistemas de control intracelular.
Nivel ontogenético. Primero necesitamos definir el concepto de “individuo”. Un individuo (individuo, individuo) es una unidad elemental indivisible de vida en la Tierra. (En algunos casos, la cuestión de determinar los límites de un individuo no es tan obvia, por ejemplo, para las colonias de pólipos y líquenes). Desde un punto de vista evolutivo, un individuo debe considerarse todas las unidades morfofisiológicas que se originan a partir de un cigoto, gametos, esporas, yemas y que están sujetas individualmente a la acción de factores evolutivos elementales.
A nivel ontogenético, la unidad de vida es el individuo desde el momento de su origen hasta la muerte. La ontogénesis es el proceso de desarrollo, implementación de información hereditaria codificada en las estructuras de control de la célula germinal. A nivel ontogenético, no solo ocurre la implementación de la información hereditaria, sino también su verificación al verificar la coherencia en la implementación de las características hereditarias y el funcionamiento de los sistemas de control en el tiempo y el espacio dentro del individuo. A través de la valoración del individuo en el proceso de selección natural se pone a prueba la viabilidad de un determinado genotipo. La ontogénesis surgió tras la adición de reduplicación convariante por nuevas etapas de desarrollo. En el curso de la evolución, el camino del genotipo al fenotipo, del gen al rasgo, surge y gradualmente se vuelve más complicado.
Las estructuras elementales en el nivel ontogenético de la organización de la vida son las células y los fenómenos elementales son algunos procesos asociados con la diferenciación.
Nivel de población-especie. La unificación de individuos en población y de poblaciones en especies según el grado de unidad genética y ecológica conduce al surgimiento de nuevas propiedades y características en la naturaleza viva, diferentes de las propiedades de los niveles genético y ontogenético molecular.
Una población es una estructura elemental a nivel de población-especie, y un fenómeno elemental a este nivel es un cambio en la composición genotípica de la población; el material elemental en este nivel son las mutaciones. Se identifican factores elementales que operan a este nivel: proceso de mutación, oleadas poblacionales, aislamiento y selección natural. Cada uno de estos factores puede ejercer una u otra “presión”.
Las poblaciones son unidades elementales y las especies son etapas cualitativas del proceso de evolución. En general, a nivel de población-especie, el proceso de evolución en realidad tiene lugar a lo largo de generaciones.
El entorno específico para el proceso de evolución que tiene lugar en poblaciones individuales es la biogeocenosis. Al mismo tiempo, la biogeocenosis es una unidad elemental del siguiente nivel de organización de la vida en la Tierra.
Nivel biogeocenótico (ecosistema). Un ecosistema es un concepto "adimensional", pero hay una clase de ecosistemas que tiene un cierto tamaño y es de fundamental importancia como "bloques de construcción" de la organización de toda la biosfera: las biogeocenosis. La biogeocenosis es un ecosistema dentro del cual no existen límites biocenóticos, microclimáticos, edáficos e hidrológicos. La biogeocenosis es uno de los sistemas naturales más complejos. Las biogeocenosis son un entorno para la evolución de sus poblaciones constituyentes. A este nivel también ocurren procesos evolutivos; la composición de la población de la biogeocenosis puede cambiar.
3. Macroevolución. Proceso evolutivo en la familia de los caballos.
La macroevolución es un proceso que se desarrolla durante un largo período de tiempo, abarca vastos territorios y conduce a la formación de nuevos grupos taxonómicos.
Género, familia, orden, clase, filo y reino representan realidades de una calidad completamente diferente a la de las especies. La integridad de los taxones de los rangos más altos no está determinada por la integración genética de sus unidades constitutivas individuales (poblaciones), como se observa dentro de una especie, sino por la unidad del "plan estructural", basado en un origen común.
El proceso evolutivo ocurre tanto en el tiempo como en el espacio. O cambian las condiciones para la existencia de la especie en el territorio que ocupa, o los cambios en las condiciones están asociados con su colonización de nuevos territorios.
Un ejemplo clásico es la historia de la especiación en la familia de los caballos (V.O. Kovalevsky). Muestra la estrecha dependencia del desarrollo histórico de este grupo de animales de los cambios ambientales.
La familia de los caballos se caracteriza por las siguientes características: las cuencas de los ojos están totalmente delimitadas por huesos; dientes con corona muy alta; su superficie de masticación está cubierta con esmalte plegado; el cúbito y el radio se fusionan y el peroné se reduce; Ambos pares de extremidades tienen un solo dedo, solo el tercer dedo está desarrollado.
Como demostró V. O. Kovalevsky, el caballo moderno de un dedo se desarrolló a partir de una forma que tenía una extremidad de cinco dedos. Los antepasados de la familia de los caballos fueron Phenacodus, que vivió en el Paleoceno. Se trata de animales relativamente pequeños con una cola larga y extremidades de cinco dedos que terminan en pequeñas pezuñas. Eran formas semidigitales, es decir, al caminar tocaban el suelo con la superficie inferior de sus falanges. Fe-nakodus tenía dientes con raíces largas y una corona baja, equipada con tubérculos a lo largo de la superficie superior. La presencia de dientes tuberosos indica que los fenacodus eran omnívoros.
Su descendiente, el representante más antiguo de la familia de los caballos Eohippus, vivió en el Eoceno inferior. Era un animal pequeño, del tamaño de un zorro, con una cabeza en la que la cuenca del ojo no estaba delimitada por la parte posterior por huesos. En sus molares hubo una desviación del tipo tuberculado, ya que en ellos aparecieron pliegues de esmalte en forma de V, lo que indica el consumo de alimentos predominantemente vegetales. El cúbito y el radio se dividieron nuevamente, pero ya había comenzado una reducción de los rayos marginales de las extremidades: las extremidades anteriores tenían cuatro dedos y las traseras solo tres. Vivía en los bosques tropicales de América del Norte y comía vegetación suculenta. De Eohippus surgió el estrechamente relacionado Orohippus, que se diferenciaba sólo en algunos cambios en la estructura de los dientes.
Al final del Eoceno, la naturaleza de la vegetación comenzó a cambiar: los cereales se generalizaron. Hacia el Mioceno, el clima cálido y húmedo da paso a uno seco y templado. En los antepasados del caballo, la selección en nuevas condiciones contribuyó a la formación de una serie de adaptaciones para alimentarse de nuevos alimentos (los dientes, la mandíbula, los músculos masticadores y los órganos digestivos cambiaron). La estructura de los órganos de movimiento cambió, lo que ayudó a escapar de los depredadores esteparios. El cuerpo del animal aumentó.
El mesohippus del Oligoceno ya tenía el tamaño de una oveja. En esta forma, la órbita en la parte posterior tampoco estaba todavía delimitada por huesos; las cuatro extremidades tenían tres dedos. Al mismo tiempo, el tercer dedo recibió el mayor desarrollo. El tipo de dientes herbívoros en Mesohippus era más pronunciado que en formas anteriores.
A partir del Mioceno se ha producido un fuerte proceso divergente que llevó a la formación de un gran número de ramas laterales que se bifurcaron a partir del tronco principal del árbol filogenético de los caballos. En el merigippus del Mioceno, los huesos del radio y el cúbito comenzaron a fusionarse, como en un caballo moderno. Los dedos marginales ya se habían acortado tanto que cuando corría el animal solo usaba un tercer dedo.
En el Plioceno, uno de los ancestros más cercanos del caballo moderno, el Pliohyppus de tres dedos, del tamaño de un burro, se trasladó a Europa junto con otros emigrantes norteamericanos. De él surgió la carrera de los caballos, los gatos. extendido por toda Eurasia y ambas Américas. En el período Cuaternario, las especies de caballos americanos se extinguieron, mientras que en Eurasia se formaron varias especies de estepas y desiertos.
Como resultado de un proceso histórico tan largo de desarrollo, surgió una forma específica de animal, que llevaba un estilo de vida gregario y pastaba en vastos espacios abiertos. En relación con la vida en estas condiciones, desarrolló una serie de adaptaciones características que, además de alimentarse de alimentos herbáceos y la velocidad de movimiento, también incluyen un embarazo prolongado y el nacimiento de cachorros, capaces de seguir a la madre poco después del nacimiento.
El proceso de formación de especies alopátricas, generalmente asociado con un cambio en el área de distribución de las especies originales. Estos cambios pueden ser de doble naturaleza: o la especie expande su área de distribución ocupando un nuevo territorio, o su área de distribución se desmembra bajo la influencia de la aparición de barreras físicas, lo que conduce al aislamiento de poblaciones individuales. En ambos casos, el camino hacia la formación de una nueva especie pasa por la aparición de nuevas subespecies.
4. Microevolución. La naturaleza del proceso microevolutivo en el ejemplo de los herrerillos, las gaviotas y las salamandras.
La microevolución es un proceso que ocurre dentro de una población y conduce a la diferenciación de la especie: la desintegración de la especie en grupos intraespecíficos de varios rangos. En el sentido moderno, el término "microevolución" fue propuesto por primera vez por Dobzhansky (1937) y Timofeev-Resovsky (1938).
El carbonero común en el Pleistoceno se dividió en tres grupos: el carbonero común, el carbonero de Bukhara y el llamado carbonero común. El primero se distingue por su mayor tamaño, dorso verde y vientre amarillo. Las tetas de Bukhara son de tamaño mediano, el color de la espalda y el vientre es gris. Las tetas “menores” son las más pequeñas, con el dorso amarillo. Después de que el hielo se retiró, estos grupos se reunieron nuevamente y sus relaciones cambiaron. El grupo de Bukhara se cruzó libremente con los otros dos, mientras que los “pequeños” y los realmente grandes, cuando se encuentran, se comportan como especies diferentes, sin cruzarse entre sí.
Estas “especies” son los eslabones finales de una única cadena de subespecies, interconectadas por zonas de integración. Para tales formas, cat. están en la frontera entre subespecies y especies, E. Mayr introdujo el término "semiespecie".
Al parecer, este fenómeno es de naturaleza muy extendida.
Gaviotas. En Europa occidental conviven dos especies de gaviotas (la gaviota argéntea y la gaviota de pico negro), el gato. fueron clasificadas como especies independientes. Existen en el mismo territorio, pero no se cruzan. Al mismo tiempo, estas formas están unidas por una serie de subespecies, formando un anillo continuo alrededor del Océano Ártico. En una compleja cadena de subespecies, estas dos formas de convivencia representan semiespecies típicas.
Salamandras. En la salamandra norteamericana Ensatina eschscholtzi, el hábitat se asemeja a una elipse, a lo largo de cuya periferia se encuentran cadenas montañosas que enmarcan su parte central de tierras bajas desérticas. La salamandra vive en la parte montañosa de su área de distribución. La especie se divide en varias razas geográficas, claramente distinguibles por el color. Las subespecies que viven cerca tienen transiciones. Pero en el sur de California la forma eschscholtzi convive con croceater y klauberi, sin cruzarse con ellos. Por tanto, estas tres formas son semiespecies.
El aislamiento de una nueva especie se producirá cuando la acción de los mecanismos de aislamiento se extienda a todas las demás poblaciones de esta especie. En otras palabras, las semiespecies son especies en proceso de convertirse, “especies emergentes”.
5. La unidad elemental del proceso evolutivo. Especie e individuo. Propiedades básicas de una población.
La unidad evolutiva deberá cumplir las siguientes condiciones:
Debe aparecer en el tiempo y el espacio como una unidad.
debe ser hereditario
Debe existir real y concretamente.
Una población es el grupo más pequeño de individuos de una misma especie que se reproducen a sí mismos, que habitan un determinado espacio durante un tiempo evolutivamente largo, forman un sistema genético independiente y forman su propio nicho ecológico.
La especie también tiene cierta unidad, pero la población es la más pequeña.
El individuo es más pequeño, pero no tiene su propio “destino evolutivo” en una serie de generaciones.
Las principales características ecológicas de la población son el tamaño, el área, la estructura por edades y sexos, así como la dinámica poblacional.
La población tiene un área específica. Los individuos fuera de este rango abandonan la población. El alcance de la población puede ampliarse, pero para ello la población debe dominar este nuevo espacio. El tamaño de la población depende en gran medida del grado de movilidad de los individuos: "los radios de la actividad individual o, más precisamente, reproductiva" (Timofeev-Resovsky). En muchos otros casos, el área trófica no coincide con el área reproductiva (desde un punto de vista evolutivo-genético, nos interesa principalmente el área reproductiva).
Relacionado con las cuestiones sobre el tamaño de las poblaciones está el problema de los números mínimos. El número mínimo es el número por debajo del cual la población inevitablemente desaparece por diversas razones ecológicas y genéticas. El tamaño de la población, al igual que otras características de la población, varía. En cada caso concreto, el tamaño mínimo de población será específico para diferentes especies.
Dinámica. El tamaño de la población (espacial y de número de individuos) está sujeto a fluctuaciones constantes. Las causas de la dinámica de las poblaciones en el espacio y el tiempo son sumamente diversas y, en términos generales, se reducen a la influencia de factores bióticos y abióticos.
Composición sexual de la población. Se sabe que el mecanismo genético de determinación del sexo asegura que la descendencia esté separada por sexo en una proporción de 1:1 (proporción primaria de sexos). Debido a la viabilidad desigual del organismo masculino y femenino (diferente viabilidad, sin duda un rasgo desarrollado evolutivamente), esta proporción primaria a veces es notablemente diferente de la secundaria (típica durante el parto en los mamíferos) y aún más notablemente diferente de la terciaria. Nogo característico para adultos.
6. La población es la unidad corogenética de evolución más pequeña. Tipos de zonas de población. La población como unidad corogenética.
Khoros – espacio, lugar, área. ……
Nekot. Las especies tienen un doble rango: trófico y reproductivo.
Las gaviotas rosadas anidan en la tundra húmeda de los ríos del noreste de Siberia. El rango trófico no ha sido determinado.
La anguila de río pasa la mayor parte de su tiempo en las aguas dulces de los ríos Negro, Azov y otros mares y lagos. Para reproducirse, migra al agua de mar (300-400 m, +7˚С). Luego, los alevines migran a los ríos.
7. El aislamiento como factor de emisión. Geográfico. Varios tipos de aislamiento biológico.
El aislamiento es la aparición de barreras que limitan la panmixia. La importancia del aislamiento en el proceso de evolución se reduce a la interrupción del libre cruce, lo que conduce a un aumento y consolidación de las diferencias entre poblaciones y partes individuales de toda la población de la especie. Sin tal consolidación de las diferencias evolutivas, no es posible la formación de formas. En la naturaleza existen: aislamiento espacial y biológico.
El aislamiento espacial puede existir de diferentes formas: las barreras de agua separan a la población de especies "terrestres" y las barreras terrestres aíslan a la población de especies acuáticas, las colinas aíslan a las poblaciones de tierras bajas y las llanuras aíslan a las poblaciones de montañas, etc. El surgimiento del aislamiento territorial, el aislamiento mecánico es explicado por la historia del desarrollo de especies en determinados territorios. En determinados casos, la principal causa del aislamiento fue el avance de los glaciares. El aislamiento espacial dentro de una especie existe en dos de sus manifestaciones: el aislamiento por cualquier barrera entre partes de la población de la especie y el aislamiento determinado por la mayor posibilidad de apareamiento de individuos que viven cerca, es decir, el aislamiento por la distancia.
El aislamiento biológico lo proporcionan dos grupos de mecanismos: los que eliminan el cruce (precopulatorio) y el aislamiento durante el cruce (poscopulatorio).
El apareamiento de formas estrechamente relacionadas se ve obstaculizado por diferencias durante la actividad sexual y la maduración de los productos reproductivos. En la naturaleza, es común el aislamiento biotópico, en el que las posibles parejas de apareamiento no se encuentran, ya que a menudo viven en lugares diferentes. Así, algunos pinzones (Fringilla coelebs) anidan en la región de Moscú en bosques tipo taiga, y otros en rodales bajos y escasos con una gran cantidad de claros. El potencial de apareamiento cruzado de individuos de estos grupos es algo limitado. Un ejemplo interesante de aislamiento biotópico son las formas intraespecíficas simpátricas en el cuco común (Cuculus canorus). En Europa habitan varias “razas biológicas” de cucos, que se diferencian por el color genéticamente determinado de sus huevos. En Europa del Este, algunos ponen huevos azules en los nidos del colirrojo común y la tarabilla común, otros ponen huevos moteados de color claro en los nidos de pequeñas aves paseriformes que tienen huevos de un color similar. El aislamiento entre estas formas de cucos se mantiene mediante la destrucción de huevos insuficientemente camuflados por parte de las especies hospedadoras. En muchas especies, la preferencia de biotopo es un mecanismo de aislamiento eficaz.
De gran importancia en el surgimiento y mantenimiento del aislamiento biológico en formas estrechamente relacionadas es el aislamiento etológico: complicaciones del apareamiento causadas por características de comportamiento. A primera vista, diferencias insignificantes en el ritual de cortejo y en el intercambio de estímulos visuales, sonoros y químicos impedirán la continuación del cortejo.
Un mecanismo de aislamiento importante que dificulta el cruce de especies estrechamente relacionadas es la aparición de diferencias morfofisiológicas en los órganos reproductivos (aislamiento morfofisiológico).
El segundo gran grupo de mecanismos de aislamiento en la naturaleza está asociado con la aparición de aislamiento después de la fertilización (aislamiento genético intrínseco), incluida la muerte de los cigotos después de la fertilización, el desarrollo de híbridos total o parcialmente estériles, así como la viabilidad reducida de los híbridos.
Lanskoy Grigory Nikolaevich historiografía nacional de la historia económica de Rusia a principios del siglo XX. Resumen de la tesis.
... fundamental ... XXsiglo V general ... imagen ... en El camino hacia el período moderno comenzó la historiografía interna de la historia económica de Rusia. XXsiglo aprobado tres ... XXsiglo estos puntos de vista se distinguieron por la mayor estabilidad, solo complementada nuevo ... Mundo historiador: XXsiglo/ ...
Bagrova Natalya Viktorovna El discurso arquitectónico-crítico como fenómeno de la cultura nacional del siglo XX especialidad 24 00 01 – teoría e historia de la cultura
Resumen de la tesis.... en nuestro vista, delineado nuevo ... en amanecer XXsiglo, que continuó en las condiciones sociales de la construcción. nuevo ... en leyes, tendencias, son comunes... Destacar tres grupos principales... formular fundamentalmentenuevo... 15 Revzin, G.I. Cuadropaz en arquitectura. Espacio y...
"un mundo sin fronteras"
DocumentoDesde los años 70 XXsiglo, primero que nada en... el cerebro en imagen manifestaciones motoras... comunicado con niños discapacitados. Aparición fundamentalmentenuevopuntos de vistaen...ver y percibir mundoen nivel personal, ... dividido condicionalmente entres grupos: ...
Los animales que se autofecundan se reproducen, en igualdad de condiciones, dos veces más rápido que los animales dioicos. ¿Por qué prevalece la dioecia en la naturaleza? Para responder a esta pregunta, se criaron artificialmente razas de lombrices intestinales. Caenorhabditis elegans, algunos de los cuales sólo practican la fertilización cruzada, otros sólo la autofecundación. Los experimentos con estos gusanos confirmaron dos hipótesis sobre los beneficios de la fertilización cruzada. Una ventaja es la limpieza más eficiente del acervo genético de mutaciones dañinas, la segunda es la acumulación acelerada de mutaciones beneficiosas, lo que ayuda a la población a adaptarse a las condiciones cambiantes.
|
|
Precio doble para machos.
¿Por qué es necesaria la reproducción sexual, por qué se necesitan los machos? Las respuestas a estas preguntas no son tan obvias como podrían parecer.
|

El famoso evolucionista John Maynard Smith llamó la atención sobre la gravedad de este problema en su libro. La evolución del sexo(1978). Maynard Smith examinó en detalle la paradoja, a la que llamó “el doble costo del sexo”. Su esencia es que, en igualdad de condiciones, la reproducción asexual (o autofecundación) es exactamente dos veces más eficaz que la fertilización cruzada con la participación de machos (ver figura). En otras palabras, los hombres son prohibitivamente caros para la población. Rechazarlos proporciona una ganancia inmediata y muy significativa en la tasa de reproducción. Sabemos que, desde el punto de vista puramente técnico, la transición de la dioica y la fecundación cruzada a la reproducción asexual o la autofecundación es bastante posible, hay muchos ejemplos de esto tanto en plantas como en animales (ver, por ejemplo: Las hembras del dragón gigante de Komodo se reproducen; sin la participación de varones, “Elementos”, 26/12/2006). Sin embargo, por alguna razón, las razas asexuales y las poblaciones de hermafroditas autofertilizantes aún no han suplantado a las que se reproducen de forma "normal", con la participación de los machos.
¿Por qué todavía son necesarios?
De lo anterior se desprende que la fertilización cruzada debe proporcionar ciertas ventajas, tan significativas que cubran incluso la doble ganancia en eficiencia reproductiva que proporciona el abandono de los machos. Además, estas ventajas deberían aparecer inmediatamente y no dentro de un millón de años. A la selección natural no le importan las perspectivas a largo plazo.
Existen muchas hipótesis sobre la naturaleza de estas ventajas (ver: Evolución de la reproducción sexual). Veremos dos de ellos. El primero se conoce como “trinquete de Muller” (ver: trinquete de Muller). Un trinquete es un dispositivo en el que el eje sólo puede girar en una dirección. La esencia de la idea es que si se produce una mutación dañina en un organismo asexual, sus descendientes ya no podrán deshacerse de él. Como una maldición familiar, se transmitirá a todos sus descendientes para siempre (a menos que se produzca una mutación inversa, y la probabilidad de que esto ocurra es muy pequeña. En los organismos asexuales, la selección sólo puede rechazar). genomas completos, pero no genes individuales). A lo largo de una serie de generaciones de organismos asexuales, puede ocurrir una acumulación constante de mutaciones dañinas (sujeto a ciertas condiciones, una de estas condiciones es un tamaño de genoma suficientemente grande en los nematodos). , Por cierto, los genomas son pequeños en comparación con los de otros animales. Quizás por eso pueden permitirse el lujo de autofertilizarse (ver más abajo).
Si los organismos se reproducen sexualmente y practican la fertilización cruzada, entonces los genomas individuales se dispersan y mezclan constantemente, y se forman nuevos genomas a partir de fragmentos que anteriormente pertenecían a diferentes organismos. Como resultado, surge una nueva entidad especial, que los organismos asexuales no tienen: reserva genética poblaciones. A los genes se les da la oportunidad de reproducirse o eliminarse independientemente unos de otros. Un gen con una mutación fallida puede rechazarse mediante selección, y los genes restantes (“buenos”) de un determinado organismo parental pueden conservarse de forma segura en la población.
Así, la primera idea es que la reproducción sexual ayuda a limpiar los genomas de la “carga genética”, es decir, ayuda a deshacerse de mutaciones dañinas que ocurren constantemente, previniendo la degeneración (una disminución en la aptitud general de la población).
La segunda idea es similar a la primera: sugiere que la reproducción sexual ayuda a los organismos a adaptarse más eficazmente a las condiciones cambiantes al acelerar la acumulación de mutaciones que son útiles en un entorno determinado. Digamos que un individuo tiene una mutación beneficiosa y otro tiene otra. Si estos organismos son asexuales, prácticamente no tienen posibilidades de esperar a que ambas mutaciones se combinen en un genoma. La reproducción sexual brinda esta oportunidad. Efectivamente convierte todas las mutaciones beneficiosas que surgen en una población en una “propiedad común”. Está claro que la tasa de adaptación a las condiciones cambiantes en organismos con reproducción sexual debería ser mayor.
Todas estas construcciones teóricas, sin embargo, se basan en ciertos supuestos. Los resultados de los modelos matemáticos indican que el grado de beneficio o daño de la fertilización cruzada en comparación con la reproducción asexual o la autofecundación depende de varios parámetros. Estos incluyen el tamaño de la población; tasa de mutación; tamaño del genoma; distribución cuantitativa de mutaciones según el grado de nocividad/utilidad; el número de crías producidas por una hembra; eficiencia de selección (el grado de dependencia del número de descendientes que quedan no de factores aleatorios, sino genéticos), etc. Algunos de estos parámetros son muy difíciles de medir no solo en poblaciones naturales, sino también en laboratorio.
Por lo tanto, todas las hipótesis de este tipo no necesitan urgentemente tanto justificaciones teóricas y modelos matemáticos (todos los cuales ya abundan), sino más bien una verificación experimental directa. Sin embargo, hasta ahora no se han llevado a cabo muchos experimentos de este tipo (Colegrave, 2002. El sexo libera el límite de velocidad de la evolución // Naturaleza. V. 420. P. 664-666; Goddard et al., 2005. El sexo aumenta la eficacia de la selección natural en poblaciones de levadura experimentales. Naturaleza. V. 434. págs. 636-640). Nueva investigación realizada por biólogos de la Universidad de Oregon sobre lombrices intestinales Caenorhabditis elegans, ilustró claramente la eficacia de ambos mecanismos considerados, proporcionando una ventaja a aquellas poblaciones que no rechazan a los machos, a pesar de su “doble precio”.
Un objeto único para estudiar el papel de los hombres
gusanos Caenorhabditis elegans como si hubieran sido creados deliberadamente para probar experimentalmente las hipótesis antes mencionadas. Estos gusanos no tienen hembras. Las poblaciones están formadas por machos y hermafroditas, siendo estos últimos numéricamente predominantes. Los hermafroditas tienen dos cromosomas X, mientras que los machos tienen solo uno (sistema de determinación del sexo X0, como Drosophila). Los hermafroditas producen espermatozoides y óvulos y pueden reproducirse sin ayuda de la autofecundación. Los machos sólo producen esperma y pueden fertilizar a los hermafroditas. Como resultado de la autofecundación, solo nacen hermafroditas. Cuando se produce la fertilización cruzada, la mitad de la descendencia son hermafroditas y la otra mitad son machos. Normalmente, la frecuencia de fertilización cruzada en poblaciones C. elegans no supera un pequeño porcentaje. Para determinar esta frecuencia, no es necesario observar la vida íntima de los gusanos; basta con conocer el porcentaje de machos en la población.
Cabe aclarar que la autofecundación no es exactamente lo mismo que la reproducción asexual (clonal), sino que las diferencias entre ellas desaparecen rápidamente en una serie de generaciones autofecundadas. Los organismos que se autofecundan se vuelven homocigotos en todos los loci a lo largo de varias generaciones. Después de esto, la descendencia deja de diferenciarse genéticamente de sus padres, de la misma forma que durante la reproducción clonal.
Ud. C. elegans Se conocen mutaciones que afectan la frecuencia de la fertilización cruzada. Uno de ellos, xol-1, es letal para los machos y, de hecho, conduce al hecho de que solo quedan hermafroditas en la población, que se reproducen por autofecundación. Otro, niebla-2, priva a los hermafroditas de la capacidad de producir esperma y los convierte efectivamente en hembras. Una población en la que todos los individuos portan esta mutación se convierte en una población dioica normal, como la mayoría de los animales.
Los autores, utilizando métodos clásicos (mediante cruces, no ingeniería genética), desarrollaron dos pares de razas de gusanos con genomas casi idénticos, que se diferencian sólo por la presencia de mutaciones. xol-1 Y niebla-2. La primera raza de cada pareja, con una mutación. xol-1, se reproduce sólo por autofecundación (autofecundación obligada, OS). En segundo lugar, con mutación. niebla-2, sólo puede reproducirse mediante fertilización cruzada (cruzamiento obligado, OO). Cada par de razas estaba acompañada por una tercera, con el mismo “antecedentes” genético, pero carente de ambas mutaciones (tipo salvaje, WT). En las razas WT, la tasa de fertilización cruzada en condiciones estándar de laboratorio no supera el 5%.
¡Se necesitan machos! Probado experimentalmente
Se llevaron a cabo dos series de experimentos con estos tripletes de rocas.
En el primer episodio Se puso a prueba la hipótesis de que la fertilización cruzada ayuda a deshacerse de la “carga genética”. El experimento continuó durante 50 generaciones (de los gusanos, por supuesto, no de los experimentadores). Cada generación de gusanos estuvo expuesta a un mutágeno químico: el metanosulfonato de etilo. Esto condujo a un aumento de aproximadamente cuatro veces en la tasa de mutación. Los animales jóvenes se colocaron en una placa de Petri dividida por la mitad por una pared de vermiculita (ver imagen), y los gusanos se colocaron en la mitad de la placa, y su alimento eran bacterias. E. coli- estaba en la otra mitad. Durante el trasplante, los gusanos fueron tratados con un antibiótico para eliminar cualquier bacteria que se hubiera adherido accidentalmente. Como resultado, para llegar a la comida y, por lo tanto, tener la oportunidad de sobrevivir y dejar descendencia, los gusanos tuvieron que superar un obstáculo. De este modo, los experimentadores aumentaron la eficacia de la selección "purificadora", que elimina mutaciones dañinas. En condiciones normales de laboratorio, la eficiencia de selección es muy baja porque los gusanos están rodeados de comida por todos lados. En tal situación, incluso los animales muy débiles y sobrecargados de mutaciones dañinas pueden sobrevivir y reproducirse. En la nueva configuración experimental, esta nivelación llegó a su fin. Para trepar por la pared, el gusano debe estar sano y fuerte.
Los autores compararon la aptitud de los gusanos antes y después del experimento, es decir, en individuos de la primera y la quincuagésima generación. gusanos C. elegans Se puede conservar congelado durante mucho tiempo. Esto facilita enormemente tales experimentos. Mientras duró el experimento, una muestra de gusanos de primera generación permaneció tranquilamente en el congelador. La aptitud se midió de la siguiente manera. Los gusanos se mezclaron en proporciones iguales con gusanos de control, en cuyo genoma se insertó el gen de la proteína brillante, y se plantaron en una configuración experimental. A los animales se les dio tiempo para superar la barrera y reproducirse, y luego se determinó el porcentaje de individuos no luminosos en la descendencia. Si este porcentaje aumentó en la quincuagésima generación en comparación con la primera, significa que la aptitud aumentó durante el experimento; si disminuyó, significa que se produjo una degeneración;
Los resultados del experimento se muestran en la figura. Indican claramente que la fertilización cruzada es un medio poderoso para combatir la carga genética. Cuanto mayor sea la frecuencia de fertilización cruzada, mejor será el resultado final (todas las líneas de la figura aumentan de izquierda a derecha). La tasa de mutación aumentada artificialmente condujo a la degeneración (disminución de la aptitud) de todas las razas de gusanos, excepto OO, "cruzadores obligados".
Incluso para aquellas razas en las que la mutagénesis no se aceleró artificialmente, la alta frecuencia de fertilización cruzada proporcionó una ventaja. En condiciones normales de laboratorio, esta ventaja no se produce porque los gusanos no tienen que trepar por las paredes para llegar a la comida.
Es curioso que en una de las dos razas de control OS ("autofertilizantes obligados"), incluso sin aumentar la tasa de mutación, el rechazo de la fertilización cruzada condujo a la degeneración (el cuadrado izquierdo en el par de curvas superiores de la figura se encuentra bajo cero).
La figura también muestra que la tasa de fertilización cruzada de la mayoría de las razas silvestres (WT) durante el experimento fue significativamente mayor que el 5% original. Este es quizás el resultado más importante. Significa que en condiciones duras (lo que significa tanto la necesidad de superar una barrera como una mayor tasa de mutagénesis), la selección natural otorga una clara ventaja a los individuos que se reproducen mediante fertilización cruzada. La descendencia de tales individuos resulta ser más viable y, por lo tanto, durante el experimento, se produce una selección por una tendencia a la fertilización cruzada.
Así, el primer experimento confirmó de manera convincente la hipótesis de que la fertilización cruzada ayuda a una población a deshacerse de mutaciones dañinas.
En el segundo episodio Los experimentos probaron si la fertilización cruzada ayuda a desarrollar nuevas adaptaciones mediante la acumulación de mutaciones beneficiosas. Esta vez, para llegar a la comida, los gusanos tuvieron que superar una zona poblada de bacterias patógenas. Serratía. Estas bacterias ingresan al tracto digestivo. C. elegans, provocan una enfermedad peligrosa en el gusano que puede provocar la muerte. Para sobrevivir en esta situación, los gusanos tuvieron que aprender a no ingerir bacterias dañinas o desarrollar resistencia a ellas. Se desconoce qué opción eligieron las poblaciones de gusanos experimentales, pero a lo largo de 40 generaciones, las razas OO se adaptaron perfectamente a las nuevas condiciones, las razas WT se adaptaron algo peor y las razas OS no se adaptaron en absoluto (su supervivencia en un entorno con condiciones nocivas). las bacterias permanecieron en el nivel bajo original). Nuevamente, durante el experimento, la frecuencia de fertilización cruzada aumentó marcadamente en las razas WT bajo la influencia de la selección.
Por tanto, la fertilización cruzada en realidad ayuda a una población a adaptarse a las condiciones cambiantes, en este caso la aparición de un patógeno. El hecho de que las tasas de fertilización cruzada aumentaran en las razas WT durante el transcurso del experimento significa que el apareamiento con machos (a diferencia de la autofecundación) da a los hermafroditas una ventaja adaptativa inmediata que aparentemente supera el "doble precio" que tienen que pagar por produciendo machos.
Cabe señalar que la fertilización cruzada ocurre no solo en organismos dioicos. Por ejemplo, muchos invertebrados son hermafroditas y no se fertilizan a sí mismos, sino que se fertilizan entre sí. En las plantas, la polinización cruzada de individuos bisexuales (“hermafroditas”) también es, por decirlo suavemente, no infrecuente. Ambas hipótesis probadas en este trabajo son bastante aplicables a estos hermafroditas. En otras palabras, este trabajo no demostró que el “hermafroditismo cruzado” sea de ninguna manera inferior a la dioicidad. Pero por la primera de estas dos opciones no es necesario pagar el famoso “doble precio”. Por tanto, el problema persiste.
Los experimentos revelaron las desventajas de la autofecundación en comparación con la fertilización cruzada, pero no explicaron por qué muchos organismos preferían la dioica a la “fertilización cruzada”. La clave para resolver este misterio probablemente resida en la selección sexual. La dioicidad permite a las hembras elegir meticulosamente a sus parejas, y esto puede servir como una forma adicional de aumentar la eficiencia para rechazar mutaciones dañinas y acumular mutaciones beneficiosas. Quizás esta hipótesis algún día reciba confirmación experimental.
Enviar su buen trabajo en la base de conocimientos es sencillo. Utilice el siguiente formulario
Los estudiantes, estudiantes de posgrado y jóvenes científicos que utilicen la base de conocimientos en sus estudios y trabajos le estarán muy agradecidos.
Publicado en http://www.allbest.ru/
Publicado en http://www.allbest.ru/
- Introducción
- Terminología
- 3. Clasificación
- 4. Anomalías de los cromosomas sexuales.
- 5. Verdadero hermafroditismo
- 6. Disgenesia gonadal mixta
- Conclusión
Introducción
El nacimiento de niños con genitales bisexuales (genitales externos masculinos y femeninos) e intersexuales (cuando el sexo del niño no queda claro en el examen) es un problema complejo no sólo de carácter médico, sino también de carácter social, sino también de problema social.
El propósito de este ensayo es cubrir el tema “Hermafroditismo”. Para lograr el objetivo, se plantearon las siguientes tareas: describir la terminología asociada a este tema, la diferenciación sexual normal, señalar sus posibles violaciones, considerar la clasificación del hermafroditismo y analizar todos sus tipos, describir el método de diagnóstico histológico del hermafroditismo.
Terminología
La intersexualidad es la presencia en un organismo dioico de características de ambos sexos, y estas características no están completamente desarrolladas, son intermedias. Las características de ambos sexos aparecen juntas en las mismas partes del cuerpo.
El hermafroditismo es un trastorno congénito del desarrollo sexual en el que los genitales externos tienen características sexuales tanto femeninas como masculinas. El hermafroditismo provoca dificultades a la hora de clasificar a un individuo como de un género determinado. Es una forma de intersexismo. Ocurre en promedio en uno de cada 2.000 recién nacidos.
El desarrollo embrionario de tal organismo se llama intersexual; comienza normalmente, pero desde cierto punto continúa como el otro sexo. Cuanto antes cambia la dirección del desarrollo de un organismo, más pronunciada es su intersexualidad.
Hay hermafroditismo verdadero y falso. El hermafroditismo verdadero (gonadal) se caracteriza por la presencia simultánea de órganos genitales masculinos y femeninos, junto con esto hay gónadas masculinas y femeninas. El verdadero hermafroditismo es mucho menos común que el falso hermafroditismo (se describen alrededor de 150 casos en toda la literatura mundial). En el verdadero hermafroditismo, los testículos y los ovarios pueden combinarse en una glándula sexual mixta o ubicarse por separado. Las características sexuales secundarias tienen elementos de ambos sexos: un timbre de voz bajo, un tipo de cuerpo mixto (bisexual) y glándulas mamarias más o menos desarrolladas. El conjunto de cromosomas (cariotipo) en el hermafroditismo verdadero suele corresponder al cariotipo femenino; con menos frecuencia hay células que contienen el conjunto de cromosomas femeninos y células que contienen el conjunto de cromosomas masculinos (el fenómeno del llamado mosaicismo).
Si los signos del verdadero hermafroditismo se extienden no solo a las gónadas, sino también a las características sexuales internas, se trata de un verdadero hermafroditismo completo.
El falso hermafroditismo (pseudohermafroditismo) ocurre cuando existe una contradicción entre las características internas (cromosómicas y gonadales) y externas (estructura de los órganos genitales) del sexo (desarrollo bisexual), es decir, las gónadas se forman correctamente según el tipo masculino o femenino, pero los genitales externos presentan signos de bisexualidad. Los factores endocrinos y los mecanismos cromosómicos juegan un papel importante en la diferenciación sexual. Las alteraciones en la relación entre las hormonas secretadas por la corteza suprarrenal, las gónadas, la glándula pituitaria de la madre y el feto y la placenta afectan la correcta formación del sexo del niño durante el período intrauterino, lo que puede provocar la presencia de gónadas de un sexo y características sexuales. inherente al otro sexo. Es casi imposible determinar el género en el pseudohermafroditismo basándose en la apariencia y estructura de los genitales externos.
El pseudohermafroditismo masculino se asocia con el tipo masculino 46 XY, que tiene características sexuales duales o genitales externos femeninos. Las anomalías pueden variar desde un simple hipospadias hasta un fenotipo femenino completo. Estos trastornos son el resultado de una estimulación androgénica insuficiente del desarrollo genital y, en la mayoría de los casos, se deben al subdesarrollo de las células de Leydig, defectos en la biosíntesis de testosterona y resistencia androgénica parcial o completa. Esto puede ser causado por tumores feminizantes raros de la corteza suprarrenal de la madre, tratamiento excesivo con hormonas sexuales femeninas, etc.
El desarrollo en las niñas de genitales externos de tipo masculino y otras características sexuales masculinas secundarias en presencia de ovarios, útero y trompas determina el falso hermafroditismo femenino. Se caracteriza por un cariotipo 46 XX, estructuras de conductos de Müller normales, ausencia de estructuras de conductos de Wolff y virilización de los genitales externos. Ocurre como resultado de un tumor en la corteza suprarrenal de la madre, tratamiento durante el embarazo con hormonas masculinas y algunos otros fármacos hormonales. El hermafroditismo masculino y femenino, a su vez, se divide en externo, interno y completo. Ante la presencia de gónadas masculinas y la similitud de los órganos genitales externos con los femeninos, se habla de falso hermafroditismo masculino externo; en presencia de testículos y al mismo tiempo útero, trompas, próstata poco desarrollada y vesículas seminales, sobre falso hermafroditismo masculino interno; una combinación de desarrollo anormal de los órganos genitales externos e internos, sobre un hermafroditismo falso completo.
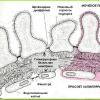
1. Diferenciación sexual normal
Etapas y mecanismos de diferenciación sexual. En el momento de la fecundación se determina el sexo genético del embrión (el conjunto de cromosomas sexuales del cigoto). El sexo genético determina la formación del sexo gonadal (formación de gónadas masculinas o femeninas). A su vez, el sexo gonadal determina la formación del sexo fenotípico (la formación de los conductos genitales y órganos genitales externos según el tipo masculino o femenino).
Diferenciación de las gónadas (formación del sexo gonadal). En la tercera semana de embriogénesis, aparecen células germinales primarias en la pared del saco vitelino, las precursoras de las oogonias y las espermatogonias. En la cuarta semana, aparecen engrosamientos en las superficies mediales de los riñones primarios: los cordones sexuales. Estos son los rudimentos de las gónadas, que consisten en células mesenquimales del riñón primario y cubiertas por epitelio celómico. Inicialmente, los cordones sexuales en los embriones masculinos y femeninos no difieren (gónadas indiferentes).
En la quinta o sexta semana de embriogénesis, las células germinales primarias pasan del saco vitelino a los cordones sexuales. Migran a lo largo de los vasos sanguíneos y el mesénquima del mesenterio del intestino posterior. A partir de este momento comienza la formación del sexo gonadal. Las células germinales primarias estimulan la proliferación y diferenciación de células mesenquimales y células epiteliales celómicas en los cordones sexuales. Como resultado, las glándulas sexuales indiferentes se convierten en testículos u ovarios y se separan de los riñones primarios. Normalmente, los cordones sexuales se diferencian en ovarios si están poblados por células germinales primarias con un cariotipo de 46, XX, y en testículos si están poblados por células con un cariotipo de 46, XY. La transformación de los cordones sexuales en testículos está determinada por el gen SRY (región Y determinante del sexo), localizado en el cromosoma Y. El gen SRY codifica un factor de desarrollo testicular. Esta proteína de unión al ADN induce la transcripción de otros genes que dirigen la diferenciación testicular.
Desarrollo testicular. En la sexta a séptima semana de embriogénesis, la corteza testicular se forma a partir del epitelio celómico del cordón sexual. Posteriormente, la capa superficial de células corticales se convierte en la túnica albugínea del testículo. Desde la capa interna de la corteza, los cordones sexuales crecen hacia el estroma mesenquimatoso de la glándula. Consisten predominantemente en células epiteliales (somáticas), entre las cuales se encuentran células germinales primarias. Los cordones reproductivos junto con el estroma mesenquimatoso forman la médula testicular. Casi desde el inicio del crecimiento de los cordones sexuales, la expresión del gen SRY aumenta en las células epiteliales. Como resultado, la corteza se degenera (solo queda la túnica albugínea) y los cordones reproductivos se convierten en túbulos seminíferos contorneados. Las células epiteliales de los cordones reproductivos se diferencian en células de Sertoli y las células mesenquimales de la médula se diferencian en células de Leydig. En la novena semana de embriogénesis, las células de Sertoli comienzan a secretar factor de regresión del conducto de Müller y las células de Leydig comienzan a secretar testosterona. Bajo la influencia de la testosterona, las células germinales primordiales de los túbulos seminíferos contorneados se diferencian en espermatogonias (esto ocurre después de la semana 22).
Desarrollo de los ovarios. En la séptima semana de embriogénesis, los ovarios se separan de los riñones primarios. Desde el epitelio celómico del cordón sexual, los cordones sexuales cortos que contienen células germinales primarias crecen profundamente hacia el estroma mesenquimatoso. Las células germinales primarias se multiplican y se convierten en oogonias. Entre el quinto y sexto mes de embriogénesis, se forman alrededor de 7 millones de oogonias. Aproximadamente el 15% de las ovogonias se convierten (sin división) en ovocitos de primer orden y el resto degenera. Los ovocitos de primer orden ingresan a la primera división de la meiosis, que se bloquea en la etapa de profase. Al mismo tiempo, se desmembran los cordones genitales y se forman los folículos primordiales. Cada folículo primordial contiene un ovocito de primer orden cubierto por una única capa de células epiteliales. Luego comienza la maduración de los folículos: se forma una membrana transparente alrededor del ovocito; Las células epiteliales crecen y forman un epitelio multicapa: una capa granular. Posteriormente, el folículo desarrolla una capa exterior formada por células mesenquimales y tejido conectivo denso. La división meiótica del ovocito de primer orden se reanuda sólo en los folículos maduros (preovulatorios) bajo la influencia de la LH. Entre las semanas 17 y 20 de la embriogénesis, finalmente se forma la estructura de los ovarios. Los folículos en diferentes etapas de maduración forman la corteza ovárica. Una niña recién nacida tiene alrededor de 1 millón de folículos. Algunos folículos sufren atresia, de modo que cuando ocurre la menarca, quedan 400.000 folículos en los ovarios. La médula está formada por tejido conectivo que contiene vasos sanguíneos y nervios.
Es probable que otros factores, aún desconocidos, también influyan en la diferenciación de las gónadas.
2. Desarrollo de los conductos reproductivos. En la cuarta semana de embriogénesis, se forman pares de conductos de Wolff (mesonéfricos) junto a los cordones sexuales del mesodermo, y en la quinta semana, se forman conductos de Müller (paramesonéfricos) laterales a ellos.
Diferenciación de conductos de Wolff. Si un testículo normal se encuentra al lado del conducto de Wolff, entre las semanas 9 y 14 se forman a partir de este conducto el epidídimo, el conducto deferente, la vesícula seminal y el conducto eyaculador. La diferenciación del conducto de Wolff es estimulada por la testosterona secretada por las células de Leydig. La testosterona no se difunde hacia el lado opuesto del embrión y, por tanto, actúa sólo en el conducto de Wolff más cercano al testículo. Si hay un ovario cerca del conducto de Wolff o si el testículo no secreta testosterona, este conducto degenera.
Diferenciación de los conductos de Müller. Si hay un testículo normal al lado del conducto de Müller, este conducto degenera entre las semanas 9 y 10 de la embriogénesis. La degeneración es causada por el factor de regresión del conducto de Müller, una glicoproteína secretada por las células de Sertoli. Si la producción o acción del factor de regresión del conducto de Müller está alterada, o si hay un ovario adyacente al conducto de Müller, entonces las trompas de Falopio, la mitad del cuerpo uterino (que luego se fusiona con la mitad opuesta) y los dos superiores -A partir de este conducto se forman los tercios de la vagina. Los ovarios no participan en la diferenciación de los conductos de Müller, por lo tanto, con la disgenesia ovárica, la formación de derivados de estos conductos no se ve afectada.
3. Desarrollo de los genitales externos. El sexo fenotípico de un recién nacido está determinado precisamente por los genitales externos. Su desarrollo ocurre simultáneamente con el desarrollo del tracto urinario y del tracto gastrointestinal distal.
En la tercera semana de embriogénesis, se forma una membrana cloacal que cubre el intestino posterior. Delante de él se forma un tubérculo genital no apareado y dos pliegues genitales en el lateral. En la sexta semana, la membrana cloacal se divide en las membranas urogenital y anal, y en la octava semana se convierte en el surco urogenital en el frente y el canal anal-rectal en la parte posterior. Los pliegues genitales se dividen en 2 pares de pliegues: los pliegues genitourinarios, situados medialmente y rodeando el surco genitourinario, y los pliegues labioescrotales, situados lateralmente. Todos estos eventos ocurren antes de la formación de las gónadas y no están regulados por hormonas. Las diferencias en los genitales externos masculinos y femeninos aparecen después de la octava semana de embriogénesis. La dirección del desarrollo de los genitales externos está determinada por las hormonas sexuales, principalmente la testosterona.
En el feto masculino, la testosterona producida en los testículos llega al tubérculo genital a través de la sangre, donde la enzima 5-alfa reductasa la convierte en dihidrotestosterona. Esta hormona actúa sobre los receptores de andrógenos y provoca un rápido crecimiento del tubérculo genital. El surco urogenital avanza, sus bordes (pliegues urogenitales) se juntan y hacia la semana 12 se forma la parte esponjosa de la uretra. Los pliegues labioescrotales se fusionan caudalmente para formar el escroto. La formación de la parte esponjosa de la uretra finaliza en el cuarto mes de embriogénesis, cuando el ectodermo del pene se invagina hacia la luz de la uretra.
En un feto femenino, los niveles de testosterona en sangre normalmente son muy bajos. Por lo tanto, los genitales externos indiferentes, formados en la octava semana de embriogénesis, posteriormente sufren solo cambios menores. El tubérculo genital se convierte en el clítoris, que puede aumentar bajo la influencia de los andrógenos no sólo en el período prenatal, sino también después del nacimiento. Los pliegues genitourinarios permanecen en el mismo lugar y forman los labios menores. Los pliegues labio-escrotales aumentan sin moverse y se convierten en los labios mayores, mientras que el surco urogenital permanece abierto, formando el vestíbulo de la vagina. La posición de la abertura externa de la uretra está determinada por la semana 14 de embriogénesis. En etapas posteriores de la embriogénesis, los andrógenos ya no pueden provocar la fusión de los pliegues labio-escrotales y el desplazamiento hacia adelante de los pliegues urogenitales.
2. Posibles trastornos de la diferenciación sexual
El principal signo de trastornos de la diferenciación sexual son los genitales externos de tipo intermedio en los recién nacidos. Estos recién nacidos requieren una evaluación urgente y un seguimiento constante, ya que pueden desarrollar una crisis de sal y un shock. Es necesario lo más rápido posible descubrir la causa de la anomalía de los órganos genitales, tranquilizar y tranquilizar a los padres y, junto con ellos, elegir para el niño el sexo de crianza que mejor se adapte a la estructura y función de los órganos genitales.
La diferenciación sexual puede verse alterada en cualquier etapa de la diferenciación sexual. Los trastornos pueden ser causados por aberraciones de los cromosomas sexuales, mutaciones de genes implicados en el desarrollo del sexo gonadal y fenotípico, así como por causas no genéticas (por ejemplo, tomar medicamentos virilizantes durante el embarazo). El sexo genético depende del cariotipo del cigoto. El cariotipo 46,XX corresponde al sexo femenino y el 46,XY al sexo masculino. Causas de los trastornos sexuales genéticos:
1. Cambios en el número o estructura de los cromosomas sexuales. Por ejemplo, las variantes clásicas del síndrome de Klinefelter (cariotipo 47,XXY) y del síndrome de Turner (cariotipo 45,X) son causadas por la falta de disyunción de los cromosomas sexuales en la meiosis durante la gametogénesis. El síndrome de Turner también puede ser causado por una deleción de uno de los cromosomas X.
2. Mosaicismo en los cromosomas sexuales (XX/XY). Este mosaicismo se encuentra en un tercio de los pacientes con hermafroditismo verdadero.
3. Mutaciones puntuales de genes en los cromosomas sexuales, por ejemplo, mutaciones del gen SRY en el cromosoma Y.
Trastornos sexuales gonadales
1. La diferenciación de las gónadas de derecha e izquierda se produce de forma independiente. Por tanto, su estructura histológica puede variar. Además, en el mismo cordón sexual se pueden formar simultáneamente diferentes glándulas sexuales. Por ejemplo, los verdaderos hermafroditas tienen un testículo y un ovario en ambos lados en forma de una sola formación (ovotestis), o hay un testículo de un lado y un ovario del otro.
2. Normalmente, el desarrollo testicular está determinado por el gen SRY, localizado en el cromosoma Y. Sin embargo, este gen se encuentra en algunos pacientes gonadales y fenotípicamente masculinos que no tienen un cromosoma Y. Es probable que en tales casos el gen SRY se transfiera al cromosoma X o a un autosoma como resultado de una translocación. Los testículos también se pueden formar en pacientes con cariotipo 46.XX que no tienen el gen SRY. Se supone que estos pacientes tienen genes mutantes que dirigen la diferenciación de los cordones sexuales hacia los testículos y no hacia los ovarios.
3. En la etapa de migración de las células germinales primarias hacia los cordones sexuales, pueden ocurrir las siguientes anomalías:
Los cariotipos de las células germinales primarias y las células somáticas de los cordones sexuales no coinciden (por ejemplo, una célula 46,XX migra a un cordón sexual formado por células 46,XY). Las células germinales primarias generalmente mueren y se forma un testículo “estéril” que no contiene células germinales. Sin embargo, hay casos en los que las células germinales primordiales sobreviven. Entonces el testículo contiene sólo células germinales con un cariotipo de 46,XX.
Lo contrario también es posible. Las células germinales primarias 46,XY pueden inducir genes que dirigen la diferenciación de células somáticas 46,XX en los cordones sexuales hacia la formación de testículos.
4. A diferencia del testículo, los ovarios pueden desdiferenciarse y convertirse en formaciones similares a cordones de tejido conectivo. Posibles causas de la desdiferenciación ovárica:
Las ovogonias no forman ovocitos de primer orden.
Los ovocitos de primer orden no son capaces de realizar división meiótica (por ejemplo, debido a aneuploidía o aberraciones cromosómicas). Probablemente así es como se desarrollan las gónadas en forma de cordón en las niñas con síndrome de Turner (cariotipos 45.X o 46.X).
Los folículos no se forman alrededor de los ovocitos.
Por tanto, para el desarrollo del ovario no sólo es necesaria la presencia de cromosomas X normales en las células del cordón sexual, sino también la presencia de ovocitos normales de primer orden.
5. Los trastornos durante el desarrollo de los genitales externos provocan atresia del ano, extrofia de la vejiga o formación de una cloaca congénita, transposición del pene y del escroto (cuando el tubérculo genital se forma caudalmente a los pliegues genitales) y agenesia de el pene. Estas anomalías suelen ser causadas por trastornos de las primeras etapas de la embriogénesis y no por trastornos del sexo genético y gonadal o de la secreción de hormonas sexuales.
6. En la misma etapa, el proceso de intususcepción del pene hacia la luz de la uretra se ve interrumpido por una insuficiencia de testosterona y dihidrotestosterona o un exceso de antagonistas de andrógenos (progesterona).
3. Clasificación
1. Trastornos de los cromosomas sexuales:
A) verdadero hermafroditismo;
B) violaciones de la formación de cromosomas sexuales y mosaicos correspondientes;
B) disgenesia gonadal atípica o mixta.
2. Pseudohermafroditismo masculino (MPH):
A) disgenesia gonadal, agonadismo;
B) feminización testicular;
B) feminización testicular incompleta;
D) defectos en la biosíntesis de testosterona;
D) persistencia del oviducto.
3. Pseudohermafroditismo femenino (FPG):
A) defectos en la síntesis de esteroides;
B) tumores en la madre que producen andrógenos;
B) influencias exógenas (medicamentos).
El primer paso diagnóstico para los genitales claramente intersexuales es el análisis cromosómico, ya que las aberraciones de los cromosomas sexuales son cuantitativamente predominantes en la génesis de las anomalías genitales. En la mayoría de los casos, esto permite asignar la observación a un grupo específico. De hecho, el 50% de los pacientes con hermafroditismo verdadero tienen un cariotipo 46-XX puro en el cultivo de linfocitos y el 80% son cromatina positiva.
Con un cariotipo puramente masculino en combinación con genitales intersexuales o femeninos, hablan con cierta cautela sobre el pseudohermafroditismo masculino. Los pacientes con un conjunto de cromosomas puramente femenino y genitales incompletos o completamente masculinos se clasifican como pseudohermafroditismo femenino. Si, con cromosomas sexuales normales, además de las anomalías genitales, existen deformidades somáticas, tras excluir las causas endocrinas, se puede suponer la presencia de un síndrome familiar o esporádico de origen desconocido.
Sin embargo, recientemente esta clasificación ha dejado de satisfacer tanto a los pacientes como a los médicos. Por un lado, el uso activo del término “hermafroditismo” ha provocado una violación de la confidencialidad de la enfermedad del paciente y frecuentes interpretaciones erróneas en el entorno del paciente. El nacimiento de un niño con una estructura anormal de los genitales externos es una prueba psicológica difícil para la familia del bebé, inevitablemente conlleva problemas sociales para los familiares y la terminología existente para esta condición (hermafrodita) sólo agrava el malestar psicológico en la familia. Por otro lado, una vez que se han establecido con precisión los mecanismos de determinadas enfermedades y las capacidades diagnósticas modernas permiten establecer un diagnóstico nosológico, es necesario cambiar la clasificación para reflejar de forma más completa la naturaleza de la afección que se ha presentado. surgido. Se propone reemplazar tanto el término "hermafroditismo", que suena ofensivo para los pacientes, como la indicación del género en el diagnóstico, es decir, hermafroditismo masculino o femenino. Se recomienda utilizar el término "alteración del desarrollo sexual" (DSD). Ver tabla en diapositiva.
En el futuro, para facilitar la comprensión del problema, recurriremos a terminología más establecida.
4. Anomalías de los cromosomas sexuales.
La condición de genitales alterados en el síndrome de Turner y el síndrome de Klinefelter no puede considerarse intersexual. La posible hipoplasia de los labios menores y la hipoplasia de los genitales masculinos con testículos pequeños en adultos en la infancia no son tan notorias como para generar dudas a la hora de establecer el sexo somático a partir del examen de los genitales externos. Con otros cariotipos esto (en particular, al nacer) tampoco causa dificultades.
Si existe un patrón en mosaico de los cromosomas sexuales, en realidad se desarrollan anomalías de los genitales externos, principalmente hipertrofia del clítoris, así como una fusión parcial de los labios menores y mayores, lo que imposibilita la determinación primaria del sexo.
5. Verdadero hermafroditismo
Se puede asumir el diagnóstico de hermafroditismo verdadero si los pacientes tienen genitales duales o indeterminados. El diagnóstico definitivo sólo puede confirmarse tras laparotomía y examen histológico de los órganos genitales internos. Es necesario diferenciar el hermafroditismo verdadero del hermafroditismo masculino falso, en el que hay tejido testicular, pero no de ovarios, y del hermafroditismo femenino falso, causado por una hiperplasia virilizante de la corteza suprarrenal, caracterizada por un fuerte aumento de la excreción de 17-cetosteroides.
El desarrollo de los órganos genitales depende del predominio de la gónada femenina o masculina, su desarrollo y actividad funcional. Existen varias combinaciones de tejido testicular y ovárico. Entonces, de un lado puede haber un testículo y del otro, un ovario; los genitales pueden estar representados en ambos lados por formaciones que son anatómicamente una combinación de tejido testicular y ovárico: ovotestis; en presencia de testículo u ovario en un lado, puede localizarse un ovotestis en el otro lado y, finalmente, puede existir un ovotestis en un lado en ausencia de tejido gonadal en el otro.
El grado de desarrollo del tejido testicular y ovárico varía significativamente hasta la aparición de los folículos maduros en los ovarios y las etapas iniciales de la espermatogénesis en los testículos. Se ha establecido que el tejido testicular, presentado en forma de testículo, tiene un efecto androgénico más pronunciado en comparación con el ovotestículo. Así, en la mayoría de los pacientes al nacer, el sexo femenino se determinó en función de la presencia de ovotestis. Externamente, los pacientes pueden tener un pene pequeño, hipospadias y pliegues escrotolabiales similares en forma al escroto, que contiene un testículo o se asemeja a los labios en ausencia de gónadas. La vagina suele estar bien desarrollada, puede desembocar en la uretra posterior, el clítoris está hipertrofiado, lo que da a los genitales un aspecto femenino. En varios pacientes, en presencia de un pene, a veces se pueden encontrar espermatozoides en el semen. La excreción urinaria de estrógenos y andrógenos se encuentra entre las normas masculinas y femeninas.
6. Disgenesia gonadal mixta
Una forma de intersexismo en la que, por un lado, se detecta una gónada disgenética (gónada-cordón), por el otro, un testículo o un tumor gonadal. En la mayoría de los casos, el desarrollo de los conductos paramesonéfricos sólo se retrasa parcialmente. En algunas observaciones, los derivados de los conductos mesonéfricos (por ejemplo, los conductos deferentes) se conservan debido al efecto pronunciado de la testosterona fetal. Síntomas. Los genitales externos se forman de manera muy diferente. Debido al descenso testicular unilateral o a la presencia de componentes tubáricos y uterinos, la formación del pliegue genital (labios mayores o mitades del escroto) puede ser asimétrica. Los síntomas generales como baja estatura y otros signos opcionales en algunos casos sugieren la presencia del síndrome de Turner.
7. Pseudohermafroditismo masculino
Cariotipo masculino indudable y anlage testicular bilateral (a excepción del agonadismo), ausencia de derivados estructurales de los conductos paramesonéfricos (a excepción del síndrome de persistencia del oviducto, cuya atribución al pseudohermafroditismo masculino es controvertida). Motivos: producción disminuida o ausente de testosterona, así como acción ineficaz de la testosterona en las células de los órganos diana con niveles normales de secreción de andrógenos. Dependiendo del grado de alteración, el espectro de anomalías de los genitales externos varía desde una mujer normal (con una vagina acortada y sin cuello uterino) hasta un hombre casi normal con hipospadias o criptorquidia.
Agonadismo. Estamos hablando de una enfermedad rara, a menudo familiar. Los pacientes tienen un cariotipo de 46, XY, genitales femeninos normales o ligeramente virilizados (grado I - II según Prader) sin útero ni trompas; en algunos casos se encuentran restos de conductos mesonéfricos. Es posible que las gónadas no sean anatómicamente visibles. Sin embargo, a juzgar por el hecho de que existen signos de la influencia de la AMH (factor X), en el período embrionario debe existir tejido testicular o células de Sertoli. Los autores creen que no detectar los testículos durante la laparotomía no es suficiente para el diagnóstico de agonadismo. Sugieren que entre el agonadismo real y la "anorquia" con genitales masculinos normales, existe un espectro de anomalías de los genitales externos dependiendo de la cantidad de tejido productor de testosterona.
Feminización testicular. En la mayoría de los casos, la feminización testicular se diagnostica durante la pubertad.
Definición. Pseudohermafroditismo masculino con cariotipo 46, XY, testículos, genitales externos femeninos normales (forma completa) o genitales intersexuales (forma incompleta) debido a la resistencia de los tejidos periféricos (órganos diana) a la testosterona y la 5alfa-dihidrotestosterona producidas en cantidades normales.
Genética. Los casos familiares son comunes. Herencia en el cromosoma X, tipo recesivo. Las formas completas e incompletas no ocurren simultáneamente en la misma familia.
Se desconoce la prevalencia, ya que existen importantes dificultades para diagnosticar las formas incompletas. Para las formas completas, la prevalencia se estima en 1:60.000; en realidad es obviamente más grande.
Defectos en la biosíntesis de testosterona. Si bien los defectos en las 21 y 11-hidroxilasas conducen al pseudohermafroditismo femenino debido al aumento de la androgenización intrauterina, existen una serie de trastornos que interfieren con la síntesis de testosterona ya en el período prenatal. Estos defectos, incluida la deficiencia de 20,22-desmolasa, nunca son tan completos como para que se formen genitales femeninos normales: en la mayoría de los casos, con un cariotipo masculino y testículos normales, el útero y la parte superior de la vagina están ausentes, y la intersexualidad Los genitales externos están claramente virilizados. El hipospadias de diversos grados es especialmente característico tanto en los testículos no descendidos como en los normalmente localizados.
Persistencia del oviducto. Estamos hablando de hombres fenotípicamente normales que a menudo tienen una violación del descenso testicular, aunque por lo demás los genitales externos no tienen nada de especial. Si tales enfermedades no se han encontrado en la familia, en la mayoría de los casos el diagnóstico se realiza durante la cirugía de hernia o criptorquidia. En este caso, se encuentra un útero hipoplásico con rudimentos tubulares (por ejemplo, en el saco herniario). Se cree que la causa de la enfermedad es una deficiencia de la hormona antimulleriana (AMH, factor X) o la resistencia de los conductos paramesonéfricos a ella.
8. Pseudohermafroditismo femenino
La virilización intrauterina de un feto de sexo genético y gonadal femenino puede ser consecuencia de tres razones conocidas:
a) virilización intrauterina por la producción de andrógenos por las glándulas suprarrenales por defectos en la síntesis de esteroides (síndrome adrenogenital);
b) producción endógena de andrógenos en el cuerpo de la madre;
c) administrar hormonas u otros medicamentos a la madre.
Pseudohermafroditismo femenino debido a defectos en la síntesis de esteroides. Estamos hablando de: a) defecto de 21-hidroxilasa sin y con síndrome de pérdida de sal; b) defecto de 11-hidroxilasa; c) un defecto de deshidrogenasa, en el que se detecta una leve virilización de un feto genéticamente femenino. En el cuadro clínico se destaca la producción insuficiente de mineralo y glucocorticoides.
Pseudohermafroditismo femenino debido a tumores productores de andrógenos en los ovarios o la corteza suprarrenal en la madre. En la mayoría de los casos, la virilización materna ya se nota durante el embarazo. Pseudohermafroditismo femenino resultante de la exposición a hormonas u otras drogas durante el embarazo. La ingesta de hormonas u otros medicamentos por parte de la madre durante el embarazo puede provocar la virilización del feto. Pseudohermafroditismo femenino en una niña cuya madre experimentó síntomas transitorios durante el embarazo que indicaban influencias androgénicas.
9. Diagnóstico histológico
Una condición indispensable para examinar a un hermafrodita es un examen histológico del tejido de ambas gónadas obtenido de su biopsia. El estado de las gónadas derecha e izquierda puede ser diferente, por lo que una biopsia de una de ellas no da idea de la estructura de ambas gónadas.
Cuando las gónadas están ubicadas en la cavidad abdominal, es posible obtener material de biopsia mediante laparoscopia y laparotomía; es más preferible la laparotomía: esto permite un examen más amplio de las gónadas y los órganos pélvicos y al mismo tiempo realizar las intervenciones quirúrgicas necesarias.
Al examinar las gónadas se evalúa el estado funcional y el grado de madurez de todas las estructuras morfológicas: gonocitos, túbulos seminíferos, folículos y tejido intersticial (células de Leydig, tejido de la teca). Esto ayuda a predecir la funcionalidad potencial de la gónada y a tomar una decisión sobre la elección del sexo del hermafrodita.
diferenciación sexual hermafroditismo cromosoma
Conclusión
El tema del hermafroditismo es relevante en nuestro tiempo, porque representa no sólo un problema médico, sino también un enorme problema social para las propias personas que padecen esta enfermedad y para su entorno más inmediato.
Las capacidades de la medicina moderna permiten a este grupo de pacientes vivir una vida plena y, en algunos casos, tener hijos. Sin embargo, este problema está completamente fuera del control de la medicina moderna. Hay muchas causas del hermafroditismo, pero existen muy pocos métodos para corregirlo, a excepción de algunas formas. Las manifestaciones del hermafroditismo son diversas y no siempre pueden detectarse de inmediato. Con el hermafroditismo, una persona experimenta dificultades con la autodeterminación sexual en un entorno social.
El diagnóstico histológico (biopsia gonadal) permite evaluar el estado funcional y morfológico de las gónadas y es uno de los principales métodos para realizar este diagnóstico. La tarea de la medicina es desarrollar métodos efectivos para diagnosticar el hermafroditismo, que permitan decir con un alto grado de probabilidad sobre el diagnóstico de un niño, y desarrollar métodos para corregir y tratar el hermafroditismo en humanos, devolviendo a la persona la oportunidad. tener hijos.
Publicado en Allbest.ru
...Documentos similares
La manifestación del hermafroditismo en humanos es la presencia de características sexuales masculinas y femeninas juntas. Análisis del hermafroditismo verdadero y falso. La esencia del dimorfismo sexual, síndrome de disgenesia gonadal, masculinización incompleta. Métodos para tratar el hermafroditismo.
prueba, agregada el 28/03/2012
Los principales signos clínicos de patología congénita. Desarrollo sexual prematuro. Hermafroditismo verdadero (gonadal). Etiología y cuadro clínico. Causas y patogenia de la criptorquidia, complicaciones y pronóstico. Síndrome adrenogenital en niñas.
presentación, añadido el 07/04/2015
Definición del síndrome de Shereshevsky-Turner (disgenesia gonadal). Consideración del cuadro clínico de un trastorno del desarrollo de las gónadas provocado por una anomalía de los cromosomas sexuales. Tratamiento con hormonas sexuales femeninas. Causas y desarrollo del síndrome de Klinefelter.
presentación, añadido el 01/06/2015
El síndrome de Turner como una forma genéticamente determinada de agenesia o disgenesia primaria de las gónadas. La incidencia de la enfermedad entre los recién nacidos y las complicaciones durante el embarazo. Patogenia de los trastornos de la formación de las gónadas, métodos de diagnóstico y tratamiento.
prueba, agregada el 27/12/2010
Clasificación del desarrollo anormal de los órganos genitales. Síndrome adrenogenital: diagnóstico y tratamiento. Anomalías del desarrollo ovárico. Disgenesia gonadal. Síndrome de ovario escleroquístico: terapia hormonal y farmacológica. Posiciones incorrectas del útero.
presentación, añadido el 23/06/2012
Factores de trastornos del desarrollo de las gónadas en el período temprano del desarrollo embrionario causados por una anomalía de los cromosomas sexuales. Síntomas clínicos de Shereevsky-Turner. Métodos de tratamiento de pacientes con esta enfermedad, métodos de diagnóstico y examen.
presentación, añadido el 21/10/2013
El concepto de enfermedades hereditarias: cambios en el número o estructura de los cromosomas. Clasificación de trastornos cromosómicos causados por cambios en los cromosomas sexuales y no sexuales. Tipos básicos de herencia. Enfermedades metabólicas y trastornos inmunológicos.
presentación, añadido el 21/11/2010
Desarrollo del aparato reproductor masculino y de los genitales externos. El proceso de formación de testículos. Malformaciones de la vesícula seminal y de la próstata. Anormalidades de la uretra. Las razones del descenso prematuro del testículo son su hipoplasia y displasia.
resumen, añadido el 19/01/2015
Síndromes cuyo desarrollo es causado por cambios en el número o estructura de los cromosomas. Frecuencia de enfermedades cromosómicas entre los recién nacidos. Síndrome de Down, síndrome de Patau, síndrome de Edwards. Anomalías de la combinación de cromosomas sexuales. Síndromes de monosomía parcial.
presentación, añadido el 06/01/2013
Formas congénitas gonadales, extragonadales y extrafetales de trastornos del desarrollo sexual. Patología cromosómica. Síndrome de Shereshevsky-Turner y agenesia gonadal “pura”. Trastornos de la segregación del cromosoma X. Síndrome de Klinefelter, masculinización incompleta.
Estábamos hablando de lombrices de tierra. Elena hizo una muy buena pregunta sobre la reproducción de los gusanos y abordó el tema del hermafroditismo biológico. Por lo cual un especial y grande GRACIAS a ella, porque hace poco conseguí 6 hermafroditas muy pequeños e interesantes en un frasco, y estaba esperando el momento para presentárselos.
El hermafroditismo, como fenómeno biológico, presupone que un individuo tiene gónadas (glándulas sexuales) femeninas y masculinas, produciendo gametos (células sexuales) tanto femeninos como masculinos. Los gametos femeninos se llaman óvulos y los gametos masculinos se llaman espermatozoides. La reproducción por hermafroditismo se clasifica como sexual, ya que en ella intervienen gametos.
El nombre del fenómeno se remonta a la mitología griega. Hermafrodita es hijo de la diosa del amor Afrodita y del dios del comercio, la magia y la razón Hermes. Era extraordinariamente hermoso. Y un día conoció a la joven ninfa Salmacis. La ninfa se enamoró tan apasionadamente y no correspondida de Hermafrodito que pidió a los dioses que los unieran para siempre. Los dioses cumplieron su oración y la pareja se fusionó en un solo ser.
El dibujo, por cierto, fue hecho con achicoria)))
En el mundo animal, el hermafroditismo es un fenómeno muy popular entre los invertebrados. Así, los tipos de celentéreos, platelmintos, anélidos y muchos moluscos se reproducen mediante hermafroditismo natural.
En las formas de vertebrados superiores, el hermafroditismo normalmente está presente en determinadas especies de peces. En anfibios, reptiles, aves y mamíferos (incluidos los humanos), el hermafroditismo se considera un fenómeno de patología (trastorno) del desarrollo embrionario.
Además, se hace una distinción entre hermafroditismo sincrónico y secuencial.
Con hermafroditismo secuencial. En un individuo sólo madura un tipo de gameto. Por ejemplo, en un pez loro, primero se activa el sistema reproductor femenino, que produce huevos, luego el pez será hembra. Después de un tiempo, el pez cambia de sexo a macho y, bajo la influencia de hormonas, produce esperma, es decir, se convierte en macho.
Cuando son sincrónicos, los óvulos y los espermatozoides se producen simultáneamente dentro de un individuo. El hermafroditismo sincrónico cruzado ocurre con mayor frecuencia, por ejemplo, en sanguijuelas y lombrices de tierra. Cuando las lombrices de tierra están listas para reproducirse, desarrollan un manguito en varios segmentos del anillo anterior llamado faja.
La faja contiene huevos. Cuando los dos gusanos se encuentran, intercambian espermatozoides, que fertilizan los óvulos dentro de la nidada. Cuando un óvulo y un espermatozoide se fusionan, se forma un cigoto o óvulo fertilizado. En total, un manguito puede contener hasta 20 huevos. El manguito se ve claramente en la foto.
Algún tiempo después de la fertilización, el manguito se desprende del gusano. Sus paredes se endurecen, el color cambia de amarillento a marrón y se forma un capullo. Del capullo emergen pequeños gusanos de aproximadamente 1 mm de longitud. Estas manguitos se forman en un gusano sexualmente maduro cada semana.
El fenómeno del hermafroditismo aumenta las posibilidades de fertilización de los individuos. Al fin y al cabo, el número mínimo de individuos para el intercambio de material genético se reduce en cualquier caso a dos. En el caso de criaturas dioicas, la probabilidad ya no es del 100%, sino sólo del 50%.
Pero aquí el coeficiente de variación de la descendencia aumenta, tendrá características más diversas, lo que significa que tendrá más posibilidades de adaptarse a las condiciones ambientales cambiantes y sobrevivir, razón por la cual en formas más complejas de animales hay machos y hembras.
Hace aproximadamente un mes obtuve pequeños hermafroditas encantadores: 6 caracoles Achatina. El día anterior soñé con al menos uno, y luego aparecieron seis. Mi colega, un matemático, los trajo y me los regaló. Fue una gran sorpresa para ella que sus caracoles pusieran una nidada de huevos de los que nacían los bebés. No podía dejarlos en el salón de biología porque los bebés necesitan cuidados diarios, así que los llevé a casa para criarlos. Cuando crezcan, se los daré a los que tengan sed.
Los caracoles solo comen lechuga; el resto de la lechuga, pepinos, manzanas y repollo son ignorados por los seis. Hoy por fin tomé una foto de los caracoles. Son criaturas nocturnas; durante el día duermen enterrándose en tierra de cocoteros. Se despiertan entre las 22 y las 23 horas y empiezan a frotarse una hoja en la rádula (lengua-rallador). Si escuchas, incluso puedes oírlos crujir.
Les doy más cáscaras trituradas. A través de las cubiertas transparentes se puede ver cómo el trozo de concha ingerido se mueve a través del tracto digestivo. Se mueven con mesura, observarlos es un placer meditativo. Al principio dispuse los caracoles en cadena, pero pronto formaron un pequeño montón. Al caracol más pequeño le encanta explorar y se mueve alrededor del frasco más que los demás. Y al caracol más grande le encanta comer.