Потенциал действия типичных атипичных кардиомиоцитов. Мембранные потенциалы кардиомиоцитов. Основные типы ионных каналов сердце
В естественных условиях клетки миокарда постоянно находятся в состоянии ритмической активности. В период диастолы мембранный потенциал покоя клеток миокарда стабилен - минус 90 мВ, его величина выше, чем в клетках водителей ритма. В клетках рабочего миокарда (предсердий, желудочков) мембранный потенциал, в интервалах между следующими друг за другом ПД, поддерживается на более или менее постоянном уровне.
Потенциал действия в клетках миокарда возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран (рисунок 3).
Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации (0 фаза), начальной быстрой реполяризации (1 фаза), переходящей в фазу медленной реполяризации (фаза плато, или 2 фаза) и фазы быстрой конечной реполяризации (3 фаза) и фазы покоя -- (4фаза).
Фаза быстрой деполяризации создается активацией быстрых потенциалозависимых натриевых каналов, обеспечивающих резкое повышение проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Мембранный потенциал уменьшается от минус 90 мВ до плюс 30 мВ, т.е. во время пика происходит изменение знака мембранного потенциала. Амплитуда потенциала действия клеток рабочего миокарда составляет 120 мВ.
При достижении мембранного потенциала плюс 30 мВ инактивируются быстрые натриевые каналы. Деполяризация мембраны вызывает активацию медленных натрий-кальциевых каналов. Поток ионов Са 2+ внутрь клетки по этим каналам приводит к развитию плато ПД (фаза 2). В период плато клетка переходит в состояние абсолютной рефрактерности.
Затем происходит активация калиевых каналов. Выходящий из клетки поток ионов К + обеспечивает быструю реполяризацию мембраны (фаза 3), во время которой медленные натрий-кальциевые каналы закрываются, что ускоряет процесс реполяризации.
Реполяризация мембраны вызывает постепенное закрывание калиевых и реактивацию натриевых каналов. В результате возбудимость миокардиальной клетки восстанавливается -- это период так называемой относительной рефрактерности.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя (фаза 4) .
Способность клеток миокарда в течение жизни человека находиться в состоянии непрерывной ритмической активности обеспечивается эффективной работой ионных насосов этих клеток. В период диастолы из клетки выводятся ионы Na + , а в клетку возвращаются ионы К + . Ионы Са 2+ , проникшие в цитоплазму, поглощаются эндоплазматической сетью.
Ухудшение кровоснабжения миокарда (ишемия) ведет к обеднению запасов АТФ и креатинфосфата в миокардиальных клетках, в результате, работа насосов нарушается, вследствие этого, уменьшается электрическая и механическая активность миокардиальных клеток.
Потенциал действия и сокращение миокарда совпадают во времени. Поступление кальция из наружной среды в клетку создает условия для регуляции силы сокращения миокарда.
Удаление кальция из межклеточного пространства приводит к разобщению процессов возбуждения и сокращения миокарда. Потенциалы действия при этом регистрируются почти в неизменном виде, но сокращения миокарда не происходит. Вещества, блокирующие вход кальция во время генерации потенциала действия, вызывают аналогичный эффект. Вещества, угнетающие кальциевый ток, уменьшают длительность фазы плато и потенциала действия и понижают способность миокарда к сокращению.
При повышении содержания кальция в межклеточной среде и при введении веществ, увеличивающих вход ионов кальция в клетку, сила сердечных сокращений увеличивается.
Соотношения между фазами ПД миокарда и величиной его возбудимости показаны на рисунке 5.
Вследствие деполяризации, мембрана кардиомиоцитов становится абсолютно рефрактерна. Период абсолютной рефрактерности у нее продолжается 0,27 с. В этот период мембрана клетки становится невосприимчивой к действию других раздражителей. Наличие длительной рефрактерной фазы препятствует развитию непрерывного укорочения (тетануса) сердечной мышцы, что привело бы к невозможности осуществления сердцем нагнетательной функции.
Фаза рефрактерности несколько короче длительности ПД миокарда желудочков, который длится около 0,3 с.
Длительность ПД предсердий - 0,1 с, столько же длиться систола предсердий.
Период абсолютной рефрактерности сменяется периодом относительной рефрактерности, во время которого сердечная мышца может ответить сокращением лишь на очень сильные раздражения. Он продолжается 0,03 с.
После периода относительной рефрактерности наступает короткий период супернормальной возбудимости, когда сердечная мышца может отвечать сокращением на подпороговые раздражения.
Сердце — мышечный орган, состоящий из четырех камер:
- правого предсердия, собирающего венозную кровь из организма;
- правого желудочка, нагнетающего венозную кровь в малый круг кровообращения — в легкие, где и происходит газообмен с атмосферным воздухом;
- левого предсердия, собирающего обогащенную кислородом кровь из легочных вен;
- левого желудочка, обеспечивающего продвижение крови ко всем органам организма.
Кардиомиоциты
Стенки предсердий и желудочков состоят из поперечно-полосатой мышечной ткани, представленной кардиомиоцитами и имеющей ряд отличий от ткани скелетных мышц. Кардиомиоциты составляют около 25% от общего числа клеток сердца и около 70% массы миокарда. В составе стенок сердца имеются фибробласты, гладкомышечные клетки сосудов, эндотелиальные и нервные клетки.
В мембране кардиомиоцитов содержатся белки, выполняющие транспортные, ферментативные и рецепторные функции. Среди последних — рецепторы гормонов, катехоламинов и других сигнальных молекул. Кардиомиоциты имеют одно или несколько ядер, множество рибосом и аппарат Гольджи. Они способны синтезировать сократительные и белковые молекулы. В этих клетках синтезируются некоторые белки, специфические для определенных стадий клеточного цикла. Однако кардиомиоциты рано теряют способность делиться и их созревание, равно как и приспособление к возрастающим нагрузкам, сопровождается увеличением массы клеток и их размеров. Причины потери клетками способности делиться остаются неясными.
Кардиомиоциты отличаются по своему строению, свойствам и функциям. Различают типичные, или сократительные, кардиомиоциты и атипичные, формирующие в сердце проводящую систему.
Типичные кардиомиоциты - сократительные клетки, образующие предсердия и желудочки.
Атипичные кардиомиоциты - клетки проводящей системы сердца, обеспечивающие возникновение возбуждения в сердце и проведение его от места возникновения к сократительным элементам предсердий и желудочков.
Абсолютное большинство кардиомиоцитов (волокон) сердечной мышцы принадлежит к рабочему миокарду, который обеспечивает . Сокращение миокарда называют, расслабление - . Имеются также атипичные кардиомиоциты и волокна сердца, функцией которых является генерация возбуждения и проведение его к сократительному миокарду предсердий и желудочков. Эти клетки и волокна формируют проводящую систему сердца.
Сердце окружено перикардом — околосердечной сумкой, отграничивающей сердце от соседних органов. Перикард состоит из фиброзного слоя и двух листков серозного перикарда. Висцеральный листок, называемый эпикардом , сращен с поверхностью сердца, а париетальный — с фиброзным слоем перикарда. Щель между этими листками заполнена серозной жидкостью, наличие которой уменьшает трение сердца с окружающими структурами. Относительно плотный наружный слой перикарда защищает сердце от перерастяжения и чрезмерного переполнения кровью. Внутренняя поверхность сердца представлена эндотелиальной выстилкой, называемой эндокардом. Между эндокардом и перикардом располагается миокард - сократительные волокна сердца.
Совокупность атипичных кардиомиоцитов, образующих узлы: синоатриальный и атриовентрикулярный, межузловые тракты Бахмана, Венкебаха и Тореля, пучки Гиса и волокона Пуркинье.
Функциями проводящей системы сердца являются генерация потенциала действия, проведение его к сократительному миокарду, инициирование сокращения и обеспечение определенной предсердий и желудочков. Возникновение возбуждения в водителе ритма осуществляется с определенным ритмом произвольно, без воздействия внешних стимулов. Это свойство клеток водителя ритма получило название .
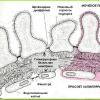
Проводящая система сердца состоит из узлов, пучков и волокон, сформированных атипичными мышечными клетками. В ее структуру входит синоатриальный (СА) узел, расположенный в стенке правого предсердия спереди устья верхней полой вены (рис. 1).

Рис. 1. Схематическое строение проводящей системы сердца
От СА-узла отходят пучки (Бахмана, Венкебаха, Тореля) атипичных волокон. Поперечный пучок (Бахмана) проводит возбуждение к миокарду правого и левого предсердий, а продольные — к атриовентрикулярному (АВ) узлу, расположенному под эндокардом правого предсердия в его нижнем углу в области, прилегающей к межпредсердной и атриовентрикулярной перегородкам. От АВ-узла отходит пучок Гпса. Он проводит возбуждение к миокарду желудочков и поскольку на границе миокарда предсердий и желудочков располагается соединительнотканная перегородка, образованная плотными фиброзными волоконами, то у здорового человека пучок Гиса является единственным путем, по которому потенциал действия может распространиться к желудочкам.
Начальная часть (ствол пучка Гиса) расположена в перепончатой части межжелудочковой перегородки и делится на правую и левую ножки пучка Гиса, которые также находятся в межжелудочковой перегородке. Левая ножка делится на переднюю и заднюю ветви, которые, как и правая ножка пучка Гиса, ветвятся и заканчиваются волокнами Пуркинье. Волокна Пуркинье расположены в субэндокардиальной области сердца и проводят потенциалы действия непосредственно к сократительному миокарду.
Механизм автоматик и проведение возбуждения по проводящей системе
Генерация потенциалов действия осуществляется в нормальных условиях специализированными клетками СА-узла, который называют водителем ритма 1-го порядка или пейсмекером. У здорового взрослого человека в нем ритмично генеририруются потенциалы действия с частотой 60-80 за 1 мин. Источником этих потенциалов являются атипичные круглые клетки СА-узла, имеющие небольшие размеры, содержащие мало органелл и редуцированный сократительный аппарат. Иногда их называют Р-клетками. В узле имеются также клетки вытянутой формы, занимающие промежуточное положение между атипичными и обычными сократительными кардиомиоцитами предсердий. Их называют переходными клетками.
Р-клетки покрыты , содержащей ряд разнообразных ионных каналов. Среди них имеются пассивные и потенциалзависимые ионные каналы. Потенциал покоя в этих клетках составляет 40-60 мВ и является неустойчивым, что обусловлено различной проницаемостью ионных каналов. Во время диастолы сердца мембрана клетки самопроизвольно медленно деполяризуется. Этот процесс назван медленной диастолической деполяризацией (МДД) (рис. 2).

Рис. 2. Потенциалы действия сократительных миоцитов миокарда (а) и атипичных клеток СА-узла (б) и их ионные токи. Пояснения в тексте
Как видно на рис. 2, сразу же после окончания предыдущего потенциала действия начинается спонтанная МДД мембраны клетки. МДД в самом начале ее развития обусловлена входом ионов Na+ через пассивные натриевые каналы и задержкой выхода ионов К+ вследствие закрытия пассивных калиевых каналов и снижения выхода ионов К+ из клетки. Вспомним, что выходящие через эти каналы ионы К обычно обеспечивают реполяризацию и даже некоторую степень гиперполяризации мембраны. Очевидно, что снижение проницаемости калиевых каналов и задержка выхода ионов К+ из Р-клетки вместе с поступлением в клетку ионов Na+ будут вести к накоплению положительных зарядов на внутренней поверхности мембраны и развитию МДД. МДД в области значений E кр (около-40 мВ) сопровождается открытием потенциалзависимых медленных кальциевых каналов, через которые в клетку поступают ионы Са 2+ , обусловливающие развитие поздней части МДД и фазы ноль потенциала действия. Хотя допускается, что в это время возможно дополнительное поступление в клетку ионов Na+ через кальциевые каналы (кальций-натриевые каналы), но решающую роль в развитии самоускоряющейся фазы деполяризации и перезарядке мембраны играют входящие в пейсмекерную клетку ионы Са 2 +. Генерация потенциала действия развивается относительно медленно, так как вход ионов Са 2+ и Na+ в клетку происходит через медленные ионные каналы.
Перезарядка мембраны ведет к инактивации кальциевых и натриевых каналов и прекращению входа ионов в клетку. К этому времени нарастает выход из клетки ионов К+ через медленные потенциалзависимые калиевые каналы, открытие которых происходит при E кр одновременно с активацией упоминавшихся кальциевых и натриевых каналов. Выходящие ионы К+ реполяризуют и несколько гиперполяризуют мембрану, после чего их выход из клетки задерживается и таким образом процесс самовозбуждения клетки повторяется. Ионное равновесие в клетке поддерживается работой натрий-калиевого насоса и натрий-кальциевого обменного механизма. Частота возникновения потенциалов действия в пейсмекере зависит от скорости спонтанной деполяризации. При возрастании этой скорости частота генерации пейсмекерных потенциалов и частота сердечных сокращений увеличиваются.
Из СА-узла потенциал распространяется со скоростью около 1 м/с в радиальном направлении на миокард правого предсердия и по специализированным проводящим путям на миокард левого предсердия и к АВ-узлу. Последний сформирован теми же типами клеток, что и СА-узел. Они также обладают способностью самовозбуждаться, но в нормальных условиях она не проявляется. Клетки АВ-узла могут начать генерировать потенциалы действия и стать водителем ритма сердца, когда к ним не поступают потенциалы действия от СА-узла. В обычных условиях потенциалы действия, возникшие в СА-узле, проводятся через область АВ-узла к волокнам пучка Гиса. Скорость их проведения в области АВ-узла резко уменьшается и промежуток времени, необходимый для распространения потенциала действия, удлиняется до 0,05 с. Эту временную задержку проведения потенциала действия в области АВ-узла называют атриовентрикулярной задержкой.
Одной из причин АВ-задержки является особенность ионных и, прежде всего кальциевых ионных, каналов мембран клеток, формирующих АВ-узел. Это находит свое отражение в более низкой скорости МДД и генерации потенциала действия этими клетками. Кроме того, клетки промежуточного участка АВ-узла характеризуются более продолжительным периодом рефрактерности, превышающим по времени фазу реполяризации потенциала действия. Проведение возбуждения в области АВ-узла предполагает его возникновение и передачу с клетки на клетку, поэтому замедление этих процессов на каждой клетке, участвующей в проведении потенциала действия, обусловливает более длительное суммарное время проведения потенциала через АВ-узел.
АВ-задержка имеет важное физиологическое значение в установлении определенной последовательности предсердий и желудочков. В нормальных условиях систола предсердий всегда предшествует систоле желудочков и систола желудочков начинается сразу же после завершения систолы предсердий. Именно благодаря АВ-задержке проведения потенциала действия и более позднего возбуждения миокарда желудочков по отношению к миокарду предсердий, желудочки заполняются необходимым объемом крови, а предсердия успевают совершить систолу (прссистолу) и изгнать дополнительный объем крови в желудочки. Объем крови в полостях желудочков, накапливаемый к началу их систолы, способствует осуществлению наиболее эффективного сокращения желудочков.
В условиях, когда нарушена функция СА-узла или имеется блокада проведения потенциала действия от СА-узла к АВ-узлу, роль водителя ритма сердца может взять на себя АВ-узел. Очевидно, что вследствие более низких скоростей МДД и развития потенциала действия клеток этого узла частота генерируемых им потенциалов действия будет ниже (около 40- 50 в 1 мин), чем частота генерации потенциалов клетками С А-узла.
Время от момента прекращения поступления потенциалов действия от водителя ритма к АВ-узлу до момента проявления его называют преавтоматической паузой. Ее длительность обычно находится в пределах 5-20 с. В это время сердце не сокращается и чем короче преавтоматическая пауза, тем лучше для больного человека.
При нарушении функции СА- и АВ-узлов водителем ритма может стать пучок Гиса. При этом максимальная частота его возбуждений составит 30-40 в 1 мин. При такой частоте сокращений сердца даже в состоянии покоя у человека будут проявляться симптомы недостаточности кровообращения. Волокна Пуркинье могут генерировать до 20 импульсов в 1 мин. Из приведенных данных видно, что в проводящей системе сердца существует градиент автомашин — постепенное снижение частоты генерации потенциалов действия ее структурами по направлению от СА-узла к волокнам Пуркинье.
Преодолев АВ-узел, потенциал действия распространяется на пучок Гиса, затем на правую ножку, левую ножку пучка Гиса и ее ветви и достигает волокон Пуркинье, где скорость его проведения возрастает до 1-4 м/с и за 0,12-0,2 с потенциал действия достигает окончаний волокон Пуркинье, с помощью которых проводящая система взаимодействует с клетками сократительного миокарда.
Волокна Пуркинье сформированы клетками, имеющими диаметр 70-80 мкм. Полагают, что это является одной из причин того, что скорость проведения потенциала действия данными клетками достигает наиболее высоких значений — 4 м/с по сравнению со скоростью в любых других клетках миокарда. Время проведения возбуждения по волокнам проводящей системы, связывающим СА- и АВ-узлы, АВ-узлу, пучку Гиса, его ножкам и волокнам Пуркинье до миокарда желудочков определяет продолжительность интервала РО на ЭКГ и колеблется в норме в пределах 0,12-0,2 с.
Не исключается, что в передаче возбуждения с волокон Пуркинье на сократительные кардиомиоциты принимают участие переходные клетки, характеризующиеся как промежуточные между клетками Пуркинье и сократительными кардиомио- цитами, структурой и свойствами.
В скелетной мышце к каждой клетке поступает потенциал действия по аксону мотонейрона и после сииаптической передачи сигнала на мембране каждого миоцита генерируется собственный потенциал действия. Взаимодействие волокон Пуркинье и миокарда совершенно иные. По всем волокнам Пуркинье к миокарду предсердий и обоих желудочков проводится потенциал действия, возникший в одном источнике — водителе ритма сердца. Этот потенциал проводится в точки контакта окончаний волокон и сократительных кардиомиоцитов в субэндокардиальной поверхности миокарда, но не к каждому миоциту. Между волокнами Пуркинье и кардиомиоцитами отсутствуют синапсы и нейромедиаторы и возбуждение может быть передано с проводящей системы на миокард через ионные каналы щелевых контактов.
Возникающий в ответ на мембранах части сократительных кардиомиоцитов потенциал проводится по поверхности мембран и по Т-трубочкам внутрь миоцитов с помощью локальных круговых токов. Потенциал передается также соседним клеткам миокарда через каналы щелевых контактов вставочных дисков. Скорость передачи потенциала действия между миоцитами достигает в миокарде желудочков 0,3-1 м/с, что способствует синхронизации сокращения кардиомиоцитов и более эффективному сокращению миокарда. Нарушение передачи потенциалов через ионные каналы щелевых контактов может быть одной из причин десинхронизации сокращения миокарда и развития слабости его сокращения.
В соответствии со строением проводящей системы потенциал действия достигает первоначально верхушечной области межжелудочковой перегородки, сосочковых мышц, верхушки миокарда. Возникшее в ответ на поступление этого потенциала в клетках сократительного миокарда возбуждение распространяется в направлениях от верхушки миокарда к его основанию и от эндокардиальной поверхности к эпикардиальной.
Функции проводящей системы
Спонтанная генерация ритмических импульсов является результатом слаженной деятельности многих клеток синусно-предсердного узла, которая обеспечивается тесными контактами (нексусы) и электротоническим взаимодействием этих клеток. Возникнув в синусно-предсердном узле, возбуждение распространяется по проводящей системе на сократительный миокард.
Возбуждение распространяется по предсердиям со скоростью 1 м/с, достигая атриовентрикулярного узла. В сердце теплокровных животных существуют специальные проводящие пути между синусно-предсердным и атриовентрикулярным узлами, а также между правым и левым предсердиями. Скорость распространения возбуждения в этих проводящих путях ненамного превосходит скорость распространения возбуждения по рабочему миокарду. В атриовентрикулярном узле благодаря небольшой толщине его мышечных волокон и особому способу их соединения (построен по принципу синапса) возникает некоторая задержка проведения возбуждения (скорость распространения составляет 0,2 м/с). Вследствие задержки возбуждение доходит до атриовентрикулярного узла и волокон Пуркинье лишь после того, как мускулатура предсердий успевает сократиться и перекачать кровь из предсердий в желудочки.
Следовательно, атриовентрикулярная задержка обеспечивает необходимую последовательность (координацию) сокращений предсердий и желудочков.
Скорость распространения возбуждение в пучке Гиса и в волокнах Пуркинье достигает 4,5-5 м/с, что в 5 раз больше скорости распространения возбуждения по рабочему миокарду. Благодаря этому клетки миокарда желудочков вовлекаются в сокращение почти одновременно, т.е. синхронно. Синхронность сокращения клеток повышает мощность миокарда и эффективность нагнетательной функции желудочков. Если бы возбуждение проводилось не через атриовентрикулярный пучок, а по клеткам рабочего миокарда, т.е. диффузно, то период асинхронного сокращения продолжался бы значительно дольше, клетки миокарда вовлекались бы в сокращение не одновременно, а постепенно и желудочки потеряли бы до 50% своей мощности. Это не позволило бы создать достаточного давления, обеспечивающего выброс крови в аорту.
Таким образом, наличие проводящей системы обеспечивает ряд важных физиологических особенностей сердца:
- спонтанную деполяризацию;
- ритмическую генерация импульсов (потенциалов действия);
- необходимую последовательность (координацию) сокращений предсердий и желудочков;
- синхронное вовлечение в процесс сокращения клеток миокарда желудочков (что увеличивает эффективность систолы).
Клетки миокарда в состоянии покоя характеризуются низкой проницаемостью для Na+, поэтому спонтанных сдвигов мембранного потенциала в них не наблюдается.
Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации (рис. 9.8). Фаза быстрой деполяризации создается резким повышением проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Происходит изменение знака мембранного потенциала с -90 до +30мВ. Деполяризация мембраны вызывает активацию медленных натрий-кальциевых каналов, в результате чего возникает дополнительный деполяризирующий входящий кальциевый ток, который приводит к фазе плато. Натриевые каналы инактивируются и клетки находятся в абсолютной рефрактерности.Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300-400 мс, что соответствует длительности сокращения миокарда. Потенциал покоя поддерживается на уровне -90мВ и определяется ионами К+.
Особенности возбудимости и сократимости миокарда.
Из материалов прошлого семестра вы помните, что возбудимость это способность возбудимой ткани под действием раздражителя переходить из состояния покоя в состояние возбуждения. Возбуждение в возбудимых тканях проявляется в виде биоэлектрических процессов и специфической ответной реакции. В сократительных клетках миокарда потенциал действия имеет особенности. Особенностью потенциала действия сократительного миокарда является наличие длительной фазы медленной реполяризации, которая обусловлена входящим током ионов Са ++ . Это приводит к тому, что длительность потенциала действия кардиомиоцитов достигает 250-300 мсек. Напомню, что длительность потенциала действия мышечных волокон скелетных мышц составляет порядка 5 мсек. Между кривой потенциала действия, кривой изменения возбудимости и кривой, отражающей изменение длины мышечного волокна существуют определенные соотношения В отличие от скелетной мышцы, у которой потенциал действия реализуется в течение латентного периода, в сократительном миокарде потенциал действия по времени совпадает с длительностью систолы и большей частью диастолы. Поскольку длительность высоковольтного пика совпадает с длительностью абсолютной рефрактерной фазы, сердце во время систолы и в течение 2/3 диастолы не может отвечать дополнительным возбуждением на какие-либо воздействия. К тому же в заключительной части диастолы возбудимость миокарда существенно снижена. Поэтому миокард в отличие от скелетной мышцы не способен к тетаническому сокращению. Эта особенность миокарда сформировалась в ходе эволюционного развития как приспособительный признак, поскольку основная функция сердца – функция биологического насоса. Эта функция может качественно выполняться только в условиях ритмических одиночных сокращений миокарда.
Таким образом, мы с вами видим, что два свойства миокарда возбудимость и сократимость связаны между собой и обусловливают важные функции сердца.
Экстрасистолы – это внеочередные по отношению к нормальному ритму сердца сокращения сердечной мышцы. Обычно экстрасистолы ощущаются пациентом как сильный сердечный толчок с провалом или замиранием после него. При прощупывании пульса в это время может быть выпадение пульсовой волны. Некоторые экстрасистолы могут возникать незаметно для больного.
Экстрасистола происходит при возникновении электрического импульса вне синусового узла. Такой импульс распространяется по сердечной мышце в период между нормальными импульсами и вызывает внеочередное сокращение сердца. Очаг возбуждения, в котором возникает внеочередной импульс, может появиться в любом месте проводящей системы сердца. К образованию подобного очага приводят как заболевания самого сердца (кардиосклероз, инфаркт миокарда, воспалительные заболевания сердечной мышцы, пороки сердца), так и болезни других органов.
ПодробностиВыделяют два типа потенциала действия (ПД): быстрый (миоциты предсердий и желудочков (0.3-1 м/с), волокна Пуркинье (1-4)) и медленный (SA-пейсмейкер 1 порядка (0.02), AV-пейсмейкер 2 порядка (0.1)).
Основные типы ионных каналов сердце:
1) Быстрые натриевые каналы (блокируем тетродотоксином) - клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье, атриовентрикулярного узла (низкая плотность).
2) Кальциевые каналы L типа (антагонисты верапамил и дилтиазем уменьшают плато, снижают силу серд. сокращения) - клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье, клетки синатриального и атриовентрикулярного узлов автоматии.
3) Калиевые каналы
а) Аномального выпрямления
(быстрая реполяризация) : клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье
б) Задержанного выпрямления
(плато) клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье, клетки синатриального и атриовентрикулярного узлов автоматии
в) формирующие I-тoк
, транзиторный выходящий ток волокон Пуркинье.
4) «Пейсмекерные» каналы, формирующие I f – входящий ток, активируемый гиперполяризацией встречаются в клетках синусного и атриовентрикулярного узла, а также в клетках волокон Пуркинье.
5) Лиганд-зависимые каналы
а) ацетилхолин-чувствительные калиевые каналы встречаются в клетках синатриального и атриовентрикулярного узлов автоматии, клетках миокарда предсердий
б) АТФ-чувствительные калиевые каналы свойственны клеткам рабочего миокарда предсердий и желудочков
в) кальций активируемые неспецефические каналы встречаются в клетках рабочего миокарда желудочков и волокнах Пуркинье.
Фазы потенциала действия.
Особенностью потенциала действия в сердечном мышце имеется выраженная фаза плато, за счет которой потенциал действия имеет такую большую длительность.
1): Фаза «плато» потенциала действия. (особенность процесса возбуждения):
ПД миокарда в желудочках сердца продолжается 300-350 мсек (в скелетной мышце 3-5 мсек) и имеет дополнительную фазу «плато».
ПД начинается с быстрой деполяризации клеточной мембраны (от - 90 мв до +30 мв), т.к. открываются быстрые Na-каналы и натрий поступает в клетку. За счет инверсии мембранного потенциала (+30 мв) быстрые Na-каналы инактивируются и ток натрия прекращается.
К этому времени происходит активация медленных Са-каналов и кальций поступает в клетку. За счет кальциевого тока деполяризация продолжается 300 мсек и (в отличие от скелетной мышцы) формируется фаза «плато». Затем медленные Са-каналы инактивируются. Быстрая реполяризация происходит за счет выхода ионов калия (К+) из клетки по многочисленным калиевым каналам.
2) Длительный рефрактерный период (особенность процесса возбуждения):
Все время пока продолжается фаза «плато» натриевые каналы остаются инактивированными. Инактивация быстрых Na-каналов делает клетку невозбудимой (фаза абсолютной рефрактерности , которая продолжается около 300 мсек).
3) Тетанус в сердечной мышце невозможен (особенность процесса сокращения):
Продолжительность абсолютного рефрактерного периода в миокарде (300 мсек) совпадает с продолжительностью сокращения (систола же6лудочков 300 мсек), поэтому во время систолы миокард невозбудим, не реагирует ни на какие дополнительные стимулы; суммация мышечных сокращений в сердце в виде тетануса невозможна! Миокард – единственная мышца в организме, которая всегда сокращается только в режиме одиночного сокращения (после сокращения всегда следует расслабление! ).
МП покоя сократительного кардиомиоцита составляет -80 (- 90) мВ.
- Быстрая начальная деполяризация (фаза 0) возникает вследствие открытия потенциалозависимых быстрых Na+-каналов, ионы Na+ быстро устремляются внутрь клетки и меняют заряд внутренней поверхности мембраны с отрицательного на положительный.
- Начальная быстрая реполяризация (фаза 1)
-- результат закрытия
Na+-каналов, входа в клетку ионов Cl- и выхода из неё ионов K+.
Последующая продолжительная фаза плато (фаза 2) -- МП некоторое время сохраняется приблизительно на одном уровне) -- результат медленного открытия потенциалозависимых Ca2+-каналов: ионы Ca2+ поступаютвнутрь клетки, равно как ионы и Na+, при этом ток ионов K+ из клетки сохраняется.
- Конечная быстрая реполяризация (фаза 3)
возникает в результате за-
крытия Ca2+-каналов на фоне продолжающегося выхода K+ из клетки
через K+-каналы.
- В фазу покоя (фаза 4) происходит восстановление МП за счёт обмена ионов Na+ на ионы K+ посредством функционирования специализированной трансмембранной системы -- Na+-К+-насоса.
Указанные процессы касаются именно рабочего кардиомиоцита. После абсолютного рефрактерного периода возникает состояние относительной рефрактерности, в котором миокард остаётся до фазы 4, т.е. до возвращения МП к исходному уровню. В период относительной рефрактерности сердечная мышца может быть возбуждена, но только в ответ на очень сильный стимул. Сердечная мышца не может, как скелетная мышца, находиться в тетаническом сокращении.
Автоматизм
-- способность пейсмейкерных клеток инициировать возбуждение спонтанно, без участия нейрогуморального контроля. Возбуждение, приводящее к сокращению сердца, возникает в специализированной проводящей системе сердца и распространяется посредством неё ко всем частям миокарда.
Проводящая система сердца.
Структуры, входящие в состав проводящей системы сердца: синусно-предсердный узел, межузловые предсердные пути, АВ-соединение (нижняя часть проводящей системы предсердий, прилегающая к АВ-узлу, собственно АВ-узел, верхняя часть пучка Хиса (Гиса), пучок Хиса (Гиса) и его ветви, система волокон Пуркинье.
Водители ритма.
Все отделы проводящей системы способны генерировать ПД с определённой частотой, определяющей в конечном итоге ЧСС, -- т.е. бытьводителем ритма. Однако синусно-предсердный узел генерирует ПД быстрее других отделов проводящей системы, и деполяризация от него распространяется в другие участки проводящей системы прежде, чем они начнут спонтанно возбуждаться. Таким образом, синусно-предсердный узел -- ведущий водитель ритма, или водитель ритма первого порядка. Частота его спонтанных разрядов определяет частоту биений сердца (в среднем 60-90 в минуту).
Градиент автоматии. В норме потенциалы первично возникают в синоатриальном узле за счет наличия клеток - водителей ритма первого порядка. Но другие отделы сердца в определенных условиях также способны генерировать нервный импульс. Это происходит при выключении синоатриального узла и при включении дополнительного раздражения.
При выключении из работы синоатриального узла наблюдается генерация нервных импульсов с частотой 50-60 раз в мин. в атриовентрикулярном узле - водителе ритма второго порядка. При нарушении в атриовентрикулярном узле при дополнительном раздражении возникает возбуждение в клетках пучка Гиса с частотой 30-40 раз в минуту - это водитель ритма третьего порядка.
Градиент автоматии - это уменьшение способности к автоматии по мере удаления от синоатриального узла, то есть от места непосредственной генерации импульсов автоматии.
Пейсмейкерные потенциалы. МП пейсмейкерных клеток после каждого ПД возвращается к пороговому уровню возбуждения. Этот потенциал, называемый препотенциалом (пейсмейкерным потенциалом ) -- триггер для следующего потенциала. На пике каждого ПД после деполяризации возникает калиевый ток, приводящий к запуску процессов реполяризации. Когда калиевый ток и выход ионов K+ уменьшаются, мембрана начинает деполяризоваться, формируя первую часть препотенциала. Открываются Ca2+ -каналы двух типов: временно открывающиеся Ca2+в-каналы и длительно действующие Ca2+ д-каналы.Кальциевый ток, идущий по Ca2+в-каналам, образует препотенциал, кальциевый ток в Ca2+д-каналах создаёт ПД.
ПД в синусно-предсердном и АВ-узлах создаются главным образом ионами Ca2+ и некоторым количеством ионов Na+. У этих потенциалов отсутствует фаза быстрой деполяризации перед фазой плато, которая имеется в других частях проводящей системы и в волокнах предсердия и желудочков.
Экстрасистола -- преждевременное (внеочередное) сокращение сердца, инициированное возбуждением, исходящим из миокарда предсердий, AВ-соединения или желудочков. Экстрасистола прерывает доминирующий (обычно синусовый) ритм. Во время экстрасистолы пациенты обычно ощущают перебои в работе сердца.
Постэкстрасистолическая потенциация. Изменение ритма сердца может воздействовать на сократимость миокарда и насосную функцию сердца без изменения длины кардиомиоцитов. Желудочковые экстрасистолы изменяют состояние миокарда таким образом, что последующие сокращения более сильны, чем нормальные предыдущие сокращения. Постэкстрасистолическая потенциация не зависит от наполнения желудочков, поскольку она может возникать в изолированной сердечной мышце в результате повышения содержания внутриклеточного Ca2+. Устойчивое увеличение сократимости может быть вызвано нанесением парных электрических стимулов на сердце, когда второй стимул следует тотчас после окончания рефрактерного периода от первого.
В состоянии покоя сердце нагнетает от 4 до 6 л крови в минуту, за день -- до 8-10 тыс. л крови. Тяжёлая работа сопровождается 4-7-кратным увеличением перекачиваемого объёма крови.
Показатели работы сердца рефлекторно изменяются в зависимости от напряжения О 2 и СО 2 в крови, от объема протекающей крови, от эмоционального состояния и физической нагрузки. Так, при физической нагрузке ударный объем может увеличиться в 2 - 3 раза, частота сокращений - в 3 - 4 раза, минутный объем кровообращения - в 4 - 5 раз.
Механизмы регуляции работы сердца:
1. Интракардиальные:
·Внутриклеточные (геторометрический и гомеометрический механизмы)
·Межклеточные механизмы
·Интракардиальные сердечные рефлексы
2. Экстракардиальные:
·Нервные
·Гуморальные
Интракардиальные механизмы в свою очередь подразделяются на миогенные (внутриклеточные), межклеточные и нервные (за счет внутрисердечной нервной системы).
·Внутриклеточные механизмы обусловлены свойствами кардиомиоцитов и лежат в основе закона Франка - Старлинга : чем больше приток крови, тем больше растягивается миокард во время диастолы, тем сильнее он сокращается во время систолы, т.е. чем больше крови поступает в желудочки, тем сильнее они потом сокращаются в систолу. Такой тип гемодинамической регуляции называется гетерометрическим . При растяжении напряжение развиваемое мышцей действительно увеличивается, но не за счет «увеличения зоны контакта актиновых и миозиновых протофибрил», а за счет увеличения вклада пассивного (эластичного) компонента в общее развиваемоемышечным волокном напряжение.Также этот механизм объясняется способностью Са2+ выходить из саркоплазматического ретикулума. Чем больше растянут саркомер, тем больше выделяется Са2+ в цитоплазму, обеспечивая большее сцепление актиновых и миозиновых нитей, и тем больше сила сокращений сердца.
Рис. Зависимость между длиной саркомера, степенью перекрытия актиновых и миозиновых нитей и развитием напряжения для препарата одиночного волокна миоцита. Объяснение в тексте. Активное напряжение при растяжении саркомера более чем на 2,2 мкм уменьшается.
Этот механизм служит для согласования систолических объёмов кровотока правой и левой половины сердца. Их систолический объём кровотока может изменяться от сокращения к сокращению. Если систолический объём левой половины во время какого-либо сокращения будет повышенным из-за значительного конечно-диастолического давления или объёма, при следующем сокращении ударный объём уменьшится и будет таким же, как и выброс правой половины сердца. Этот механизм саморегуляции включается при перемене положения тела, при резком увеличении объема циркулирующей крови (при переливании), а также при фармакологической блокад симпатической нервной системы бета-симпатолитиками.
·Гомеометрическая внутриклеточная регуляция сердца (феномен Анрепа и хронотропная зависимость Боудича)
Гомеометрический механизм не зависит от исходной длины кардиомиоцитов. Сила сердечных сокращений может возрастать при увеличении частоты сокращений сердца. Чем чаще оно сокращается, тем выше амплитуда его сокращений («лестница» Боудича ), однако при повышении ЧСС более 180 уд.мин сила сокращений снижается. Сердце человека и большинства животных, за исключением крыс в ответ на повышение ритма реагирует увеличением силы сокращений и, наоборот, с уменьшением ритма сила сокращений падает. Механизм этого феномена связан с накоплением или падением в миоплазме концентрации Са2 +, а следовательно - увеличением или уменьшением количества поперечных мостиков. При частом раздражении происходит увеличение ионов кальция в цитозоле, поскольку все больше ионов освобождаются из саркоплазматического ретикулума с каждым последующим мышечным потенциалом действия, а убрать их из саркоплазмы немедленно не удается,т.к. это активный, а значит медленный процесс.

Рис. Возникновение «лестницы Боудича» при увеличении частоты следования импульсов. S - стимулы, одинаковые по силе, но разные по частоте (А - более редкие, Б - более частые). R - ответы (сокращения миокард) (А - одинаковые по амплитуде, Б - возрастающей амплитуды).
При повышении давления в аорте до определенных пределов возрастает противонагрузка на сердце, происходит увеличение силы сердечных сокращений (феномен Анрепа ), обеспечивая тем самым возможность выброса того же объема крови, что и при исходной величине артериального давления, т.е. чем больше противонагрузка, тем больше сила сокращений. Механизмы, лежащие в основе феномена Анрепа, до сих пор не раскрыты. Предполагают, что с увеличением противонагрузки растет концентрация Са2+ в межфибриллярном пространстве и поэтому возрастает сила сердечных сокращений.
·Регуляция межклеточных взаимодействий. Установлено, что вставочные диски, соединяющие клетки миокарда, имеют различную структуру. Одни участки вставочных дисков выполняют чисто механическую функцию, другие обеспечивают транспорт через мембрану кардиомиоцита необходимых ему веществ, третьи -- нексусы, или тесные контакты, проводят возбуждение с клетки на клетку. Нарушение межклеточных взаимодействий приводит к асинхронному возбуждению клеток миокарда и появлению сердечных аритмий.
К межклеточным взаимодействиям следует отнести и взаимоотношения кардиомиоцитов с соединительнотканными клетками миокарда. Последние представляют собой не просто механическую опорную структуру. Они поставляют для сократительных клеток миокарда ряд сложных высокомолекулярных продуктов, необходимых для поддержания структуры и функции сократительных клеток. Подобный тип межклеточных взаимодействий получил название креаторных связей (Г. И. Косицкий).
· Внутрисердечные периферические рефлексы .
Более высокий уровень внутриорганной регуляции деятельности сердца представлен внутрисердечными нервными механизмами. Обнаружено, что в сердце возникают так называемые периферические рефлексы, дуга которых замыкается не в ЦНС, а в интрамуральных ганглиях миокарда. После гомотрансплантации сердца теплокровных животных и дегенерации всех нервных элементов экстракардиального происхождения в сердце сохраняется и функционирует внутриорганная нервная система, организованная по рефлекторному принципу. Эта система включает афферентные нейроны, дендриты которых образуют рецепторы растяжения на волокнах миокарда и венечных (коронарных) сосудах, вставочные и эфферентные нейроны. Аксоны последних иннервируют миокард и гладкие мышцы коронарных сосудов. Указанные нейроны соединяются между собой синаптическими связями, образуя внутри-сердечные рефлекторные дуги.
В экспериментах показано, что увеличение растяжения миокарда правого предсердия (в естественных условиях оно возникает при увеличении притока крови к сердцу) приводит к усилению сокращений миокарда левого желудочка. Таким образом, усиливаются сокращения не только того отдела сердца, миокард которого непосредственно растягивается притекающей кровью, но и других отделов, чтобы «освободить место» притекающей крови и ускорить выброс ее в артериальную систему. Доказано, что эти реакции осуществляются с помощью внутрисердечных периферических рефлексов (Г. И. Косицкий).
Подобные реакции наблюдаются лишь на фоне низкого исходного кровенаполнения сердца и незначительной величины давления крови в устье аорты и коронарных сосудах. Если камеры сердца переполнены кровью и давление в устье аорты и коронарных сосудах высокое, то растяжение венозных приемников в сердце угнетает сократительную активность миокарда, в аорту выбрасывается меньшее количество крови, а приток крови из вен затрудняется. Подобные реакции играют важную роль в регуляции кровообращения, обеспечивая стабильность кровенаполнения артериальной системы.
Гетерометрический и гомеометрический механизмы регуляции силы сокращения миокарда могут привести лишь к резкому увеличению энергии сердечного сокращения в случае внезапного повышения притока крови из вен или повышения артериального давления. Казалось бы, что при этом артериальная система не защищена от губительных для нее внезапных мощных ударов крови. В действительности же таких ударов не возникает благодаря защитной роли, осуществляемой рефлексами внутрисердечной нервной системы.
Переполнение камер сердца притекающей кровью (равно как и значительное повышение давления крови в устье аорты, коронарных сосудов) вызывает снижение силы сокращений миокарда посредством внутрисердечных периферических рефлексов. Сердце при этом выбрасывает в артерии в момент систолы меньшее, чем в норме, количество содержащейся в желудочках крови. Задержка даже небольшого дополнительного объема крови в камерах сердца повышает диастолическое давление в его полостях, что вызывает снижение притока венозной крови к сердцу. Излишний объем крови, который при внезапном выбросе его в артерии мог бы вызвать пагубные последствия, задерживается в венозной системе.
Опасность для организма представляло бы и уменьшение сердечного выброса, что могло бы вызвать критическое падение артериального давления. Такую опасность также предупреждают регуляторные реакции внутрисердечной системы.
Недостаточное наполнение кровью камер сердца и коронарного русла вызывает усиление сокращений миокарда посредством внутрисердечных рефлексов. При этом желудочки в момент систолы выбрасывают в аорту большее, чем в норме, количество содержащейся в них крови. Это и предотвращает опасность недостаточного наполнения кровью артериальной системы. К моменту расслабления желудочки содержат меньшее, чем в норме, количество крови, что способствует усилению притока венозной крови к сердцу.
В естественных условиях внутрисердечная нервная система не является автономной. Она -- лишь низшее звено сложной иерархии нервных механизмов, регулирующих деятельность сердца. Следующим, более высоким звеном этой иерархии являются сигналы, поступающие по блуждающим и симпатическим нервам, осуществляющие процессы экстракардиальной нервной регуляции сердца.
Эффекты воздействий на сердце:
·влияние на ЧСС (т.е. на автоматизм) обозначают термином «хронотропноедействие» (может быть положительным и отрицательным),
·на силу сокращений (т.е. на сократимость) -- «инотропное действие»
·на скорость предсердно- желудочкового проведения (что отражает функциюпроводимости) -- «дромотропное действие» (положительное или отрицательное),
·на возбудимость -- «батмотропное действие» (также положительное илиотрицательное).